

Глава 2. Сенсорная функция мозга
69
чи отражением более общего принципа конструкции центральных связей, а именно: принципа «нервной решетки». В мозговых структурах благодаря умножению числа нейронов предпосылок для широкого пространственного распространения нервных влияний гораздо больше, чем на периферии. Одновременно с этим действует принцип общего пути Ч. С. Шеррингтона, то есть схождения многих входов на одном нейроне как на «воронке». По мере восхождения от одного этажа мозга к другому «воронка» все более расширяется за счет присоединения все новых входов, и возрастают требования к все более точному выведению на «общий путь» биологически значимой информации. Ч. Шеррингтон писал по этому поводу: «...Если два или более афферентных нейрона конвергируют на третьем, который по отношению к ним является эфферентным (исполнительным. — А. С.) и на котором, как показано выше, сливаются или интерферируют их влияния, то такая структура представляет собой механизм координации» (Шеррингтон, 1969).1
Итак, наличие в сенсорных системах ряда уровней, каждый из которых работает по принципу дивергенции и конвергенции — это наличие важнейших координационных аппаратов, где происходит поэтапная обработка информации. По-видимому, оба механизма необходимы для неискаженной передачи сведений об отдельных признаках и деталях наряду с объединением их при формировании целостного образа.
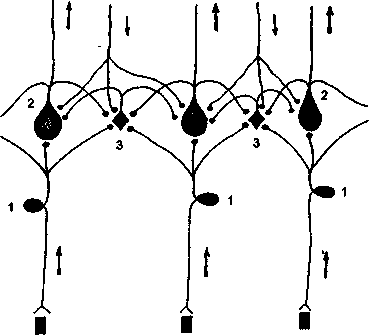
Принцип обратных связей. Деятельность сенсорной системы должна рассматриваться не как процесс пассивного кодирования любого раздражения в частотно модулированную импульсную сигнализацию, а как процесс активного восприятия и обработки наиболее биологически существенной информации. Сенсорную систему необходимо рассматривать не просто как пассивный канал линии связи, а как активный «сигнализатор». Сенсорная система состоит не из лестницы релейных образований, а из аппаратов управления процессом обработки и передачи информации с нижележащих уровней на вышележащие. И каждый уровень системы должен работать минимум на основе двух входов: 1) входа информации — восходящий путь и 2) входа управления — нисходящий путь. Таким образом, центральная организация сенсорной системы в совокупности прямых и обратных связей представляется в виде системы образований из надстраивающихся нервных колец (рис. 3).
Наличие нисходящих связей к различным образованиям сенсорной системы говорит о том, что на их работу влияют вышерасположенные отделы многих мозговых структур. Благодаря деятельности нисходящих связей обеспечивается регуляция пороговой чувствительности и пропускной способности в сенсорных системах. Это естественно, ибо сенсорная система может обработать за единицу времени объем информации во много раз меньший, чем тот, который может поступать на вход сенсорной системы.
Направленность нисходящих влияний определяется доминирующей потребностью как предпосылкой образования доминанты при использовании аппаратов памяти. В связи с этим особенно важной оказывается кортикальная регуляция, которая осуществляет три функции: 1) пусковую, 2) корригирующую и 3) поддерживающую. Значение первой из них состоит в открытии или блокировании сенсорных входов; второй — в трансформации
Рис. 3 Схема построения
нейронных
переключений
в сенсорной системе
(по Г. И. Полякову,
1965)
Периферический чувствительный нейрон, передающий центростремительные импульсы как на эфферентные нейроны с длинным аксоном, так и на специальные переключательные нейроны с коротким аксоном. Наряду с восходящими проводниками (стрелки вверх) показаны нисходящие центробежные проводники (стрелки вниз).
1 — периферический чувствительный нейрон, 2 — эфферентные нейроны с длинным аксоном, 3 — переключательные нейроны с коротким аксоном
нервной сигнализации или в обеспечении условий, при которых ранее разбалансированные сенсорные системы могут быть приведены к такому режиму деятельности, который создает необходимые условия для их наиболее полного взаимодействия и интеграции, и третий — в поддержании возбудимости глубоких структур данной сенсорной системы на уровне, оптимально приспособленном к восприятию и передаче сигнала в восходящем направлении.
Имеющиеся данные свидетельствуют о тормозном значении обратных связей. Это становится понятным, если принять во внимание чрезвычайно высокую (можно сказать, критическую) чувствительность рецепторных образований сенсорных систем. Именно на этих уровнях сенсорных систем начинается процесс активного восприятия и обработки сигналов.
Принцип кортикализации. Процесс формирования новой коры связан с представительством всей совокупности сенсорных систем. Это определило принцип функциональной многозначности коры, согласно которому все корковые области — это корреляционные центры, среди которых нет чисто проекционных центров, а значит, нет места абсолютной локализации.
Экранная структура новой коры явилась субстратом для объединения деятельности многих сенсорных систем.
Одной из общих черт корковой проекции сенсорных систем является их множественный характер представительства в коре. В самой общей форме разграничивают первичные и вторичные проекции. Первичные корковые проекции возникают в онтогенезе человека сравнительно рано, здесь заканчиваются быстропроводящие сенсорные каналы. Эти корковые зоны окружены вторичными зонами той же сенсорной системы, импульсация к которым поступает несколько позднее, чем к первичным зонам. Эти корковые поля принимают интегрированную информацию в результате взаимодействия специализированных каналов данной сенсорной системы.
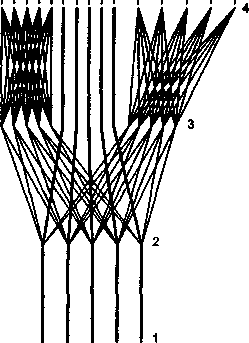
Наконец, выделяют зоны перекрытия разных сенсорных систем, где происходит межсенсорное взаимодействие (рис. 4). Эти зоны получили название третичных или ассоциативных полей. Например, в зрительной системе повреждение первичной проекционной зоны приводит к возникновению «физиологической слепоты» — исчезает восприятие противоположной половины поля зрения (гемианопсия). Повреждение же вторичных проекционных зон коры вызывает «психическую слепоту», которая именуется зрительной агнозией (неузнавание предметов). Поэтому высшим отделом сенсорной системы (в частности, зрительной) считают именно вторичные сенсорные поля, оставив за первичными — в значительной мере релейную, переключающую функцию.
Принцип двусторонней симметрии. Любая сенсорная система построена по принципу билатеральной симметрии. Иными словами, рецепторные аппараты, соответствующие им центральные проводники и центральные мозговые структуры парные, и одна половина зеркально повторяет другую. Однако, как правило, этот принцип проявляется лишь в относительной степени, ибо даже первичный сенсорный путь может быть связан с обоими симметричными мозговыми полушариями. Связь с контралатеральным полушарием выражена сильнее благодаря большему числу направляющихся туда сенсорных волокон. Между симметричными отделами сенсорных систем устанавливаются горизонтальные комиссуральные связи, обеспечивающие их взаимодействие.
Рис. 4
Схема, изображающая
обособление в корковой
сенсорной зоне центрального
поля, в котором
концентрируются прямые
проекции «точка в точку»
и периферические поля,
в которые собираются
преимущественно непрямые,
перекрестные связи
(третичные или
ассоциативные поля)
(по Г. И. Полякову, 1965)
1 — уровень рецепторов; 2 — уровень
переключений в стволе мозга; 3 —
в больших полушариях; 4 — в коре
мозга.
Основным механизмом парной деятельности сенсорной системы является механизм функциональной асимметрии при действии различным образом локализованных в пространстве объектов. Парная деятельность сенсорных систем заключается в сравнении пространственной модели ранее действовавшего стимула с новой пространственной локализацией того же стимула. Межполушарная асимметрия в пределах одной сенсорной системы функционирует как саморегулирующаяся система с обратной тормозной связью, осуществляя функцию своеобразного компаратора, выполняющего сравнение рисунков возбуждения при раздражении симметричных рецепторов. Система калловальных связей между ассоциативными полями обеспечивает тесное единство обеих половин ассоциативной системы и создает высокую надежность ее функционирования.
Принцип структурно-функциональных корреляций. Неодинаковая степень кортикализации сенсорных систем и большее развитие глубоких структур слуховой системы по сравнению со зрительной свидетельствует о том, что зрительная сигнализация для некоторых животных обладает наибольшей физиологической силой, и степень кортикализации зрительной системы настолько высока, что любое повреждение коры прежде всего сказывается на дефектах зрительного распознавания, в меньшей степени отражаясь на анализе звуковых сигналов.
У таких животных как дельфины, летучие мыши, слуховая система выполняет более важную роль в пространственной ориентации, чем зрительная система.
Любая психофизиологическая функция зависит от одновременной работы нескольких сенсорных систем, то есть является полисенсорной и поэтому не может быть локализованной в ограниченных отделах мозга. Сама же сенсорная система выступает как определенная локализованная анатомическая система, выполняющая специализированную функцию обнаружения и преобразования информации в нервный код, в котором заключена совокупность описания признаков воспринимаемого объекта или явления.