

Формы индивидуального обучения
Генетически детерминированные формы поведения, отражающие накопленный в генофонде видовой опыт предшествующих поколений, оказываются недостаточными, чтобы обеспечить активное существование особи в вероятностно изменчивой среде. Чем меньше выражен диапазон колебаний факторов среды, тем больше поведение животного может опираться на опыт предшествующих поколений, записанный в его генотипе. И наоборот, чем больше изменчивость окружающей действительности, тем меньше прагматическая ценность видового опыта, тем в большей мере возрастает необходимость в приобретении собственного, индивидуального опыта. Последний приобретается различными путями, в основе которых лежит общая способность живых организмов к обучению, которое тесно увязано со свойством фиксировать на какой-то срок элементы обучения, то есть он оснащен памятью. Поэтому нейробиология обучения и памяти — это единая проблема для науки.
Обучение — это процесс, состоящий в появлении адаптивных изменений индивидуального поведения в результате приобретения опыта (В. Торп, 1963).
Следовательно, при обучении происходит отбор оптимальных реакций животного, но сам процесс обучения, скрытый от непосредственного измерения, может быть оценен только по результатам поведения. Иными словами, мы оцениваем лишь то, что животные запомнили в результате обучения. Вероятно, не существует единых общих законов обучения, а эта важнейшая способность живых организмов должна эволюционировать как в онто-, так и в филогенетическом отношении, чтобы обеспечить поведению его адаптивный характер.
Классификация форм обучения. Обучение является предметом изучения не только физиологов, но и зоопсихологов, этологов, бихевиористов. Отсюда необычайная пестрота взглядов, противоречия отдельных школ, трудно сопоставимые методические подходы.
Рассмотрим основные категории обучения животных (табл.).
I. Молодые организмы обеспечены достаточно стабильным набором внешних стимулов, среди которых им приходится оперировать на самых ранних этапах самостоятельного Существования. Обучение в этот период носит неассоциативный, облигатный характер, в значительной степени обусловленный набором средовых факторов и не требующий непременного совпадения (ассоциации) внешних сигналов с той или иной целостной деятельностью организма, то есть является стимул-зависимым.
Таблица. Классификация обучения
Категория |
Основные формы |
Неассоциативное, облигатное, стимул-зависимое обучение |
Суммационная реакция |
Привыкание |
|
Запечатление (импринтинг) |
|
Подражание |
|
Ассоциативное, факультативное, эффект-зависимое обучение |
Классический условный рефлекс |
Инструментальный условный рефлекс (включая обучение по типу проб и ошибок) |
|
Когнитивное обучение |
Психонервная деятельность |
Рассудочная деятельность |
|
Вероятностное прогнозирование |
II. На более поздних этапах онтогенеза по мере созревания организма поведение все в большей мере приобретает активный характер. Значительно расширяется спектр внешних факторов, могущих приобретать то или иное сигнальное значение в зависимости от ассоциации их с целостной биологической реакцией организма (ассоциативное, факультативное обучение). От относительно пассивного восприятия среды организм переходит к активному процессу формирования собственной среды путем извлечения для себя ее функциональных составляющих, значимых для выполнения тех или иных актов поведения. Обучение в этот период носит эффект-зависимый характер, то есть определяемый результативностью контакта организма со средой.
III. Высшие формы обучения, свойственные в большей степени взрослым животным с высокоразвитой нервной системой, опираются на свойство формировать функциональную структуру окружающей среды в концептуальном плане, иными словами, формировать целостный образ окружающей среды. Такие формы когнитивного обучения основаны на формировании функциональной структуры среды, то есть на извлечении законов связей между отдельными ее компонентами. Естественно, чта такие категории обучения базируются на двух предыдущих и должны рассматриваться как продукт их прогрессивного развития в возрастном и филогенетическом плане.
Иногда весьма затруднительно отнести ту или иную поведенческую адаптацию к определенной категории обучения. Тем не менее богатство и разнообразие форм индивидуального обучения живых организмов может быть ранжировано исходя из эволюционных общебиологических закономерностей.
Рассмотрим последовательно основные формы обучения.
НЕАССОЦИАТИВНОЕ ОБУЧЕНИЕ
Суммационная реакция. Феномен суммации в центральной нервной системе известен давно и детально изучен как на клеточном, так и на системном уровнях организации. В основе этого явления лежит сенсибилизация — повышение чувствительности нервной ткани к раздражающим агентам и фасилитация — облегчение запуска именно данной реакции. Рядом ученых наблюдались на простейших и низших беспозвоночных реакции, как будто бы напоминающие условные рефлексы: освоение определенного маршрута передвижения, навыки скопления в определенных местах пространства, различение съедобных продуктов от несъедобных, осуществление защитных двигательных реакций в ответ на прежний индифферентный раздражитель.
Однако при углубленном анализе стало ясно, что эти навыки обладают всеми свойствами суммационной реакции: 1) они не могут сохраняться длительное время и обречены на исчезновение; 2) вызывающие их раздражители не обладают специализированным сигнальным значением; 3) после исчезновения они самостоятельно не восстанавливаются; 4) их исчезновение есть процесс разрушения, а не временного торможения.
Возникнув у одноклеточных организмов, суммационная реакция, основанная на свойствах протоплазмы к сенсибилизации и фасилитации ответных реакций, может быть рассмотрена в качестве одной из форм индивидуального приспособления у примитивных живых существ. Однако в преобразованном, а иногда тщательно замаскированном виде Суммационная реакция у высших животных выступает как важнейший элемент более сложных форм индивидуального обучения.
Клеточные механизмы суммации участвуют к процессах обучения и памяти у высших животных.
Привыкание. В противоположность суммационной реакции привыкание представляет собой такую форму обучения, которая состоит в относительно устойчивом ослаблении реакции вследствие многократного предъявления раздражителя, не сопровождающегося каким-либо биологически значимым агентом (пищевым, оборонительным, сексуальным и пр.), именуемым обычно подкреплением. Явление привыкания весьма широко распространено в животном мире, начиная с одноклеточных организмов. Привыкание у одноклеточных — это кратковременный процесс, не обладающий способностью к тренировке. У низших кишечнополостных с диффузно организованной нервной системой (у гидр) выработка привыкания в каждом последующем опыте проходила быстрее, чего не наблюдалось у инфузорий. Следовательно, у гидр, как и у инфузорий, привыкание лежит в основе физиологических механизмов, определяющих способность к приобретению индивидуального опыта (Н. А. Тушмалова, 1986).
Быть может, для столь низкоорганизованных живых форм, как одноклеточные и животные с примитивной нервной системой, единственными механизмами приобретения индивидуального опыта являются суммационная реакция и относительно кратковременное привыкание. Таким образом, повышенная реактивность к одним сигналам вместе со сниженной реактивностью к другим, сохраняющиеся на протяжении относительно краткого промежутка времени, могут выступать в качестве главных (если не единственных) механизмов адаптивных изменений индивидуального поведения, полностью зависимого от физико-химических параметров окружающей среды. Поэтому такое поведение можно именовать стимул-зависимым поведением. Привыкание усовершенствуется в ходе прогрессивной эволюции и сохраняет за собой важную роль в репертуаре поведенческих адаптации. Причем привыкание возникает не в результате утомления или сенсорной адаптации. В некотором смысле привыкание — это подавление реакций, которые оказываются незначимыми в жизни животного.
Привыкание рассматривают как простейшую форму обучения, которая может служить основой для развития более сложных форм обучения.
Дальнейшими исследованиями было установлено, что привыкание, присутствующее в самых разнообразных рефлекторных системах, не является утомлением и зависит от специфичности стимула, силы раздражения и состояния организма. Различают кратковременные и долговременные процессы привыкания, которые используются как модель для изучения разных видов памяти.
Наиболее же распространенной формой привыкания является ориентировочный рефлекс, который при повторении вызвавшего его раздражителя постепенно угашается. Раздражители, которые не имеют для животного прямого биологического значения, постепенно утрачивают ответную реакцию организма. Такая форма поведенческой адаптации, в основе которой лежит научение не отвечать на раздражение (негативное научение), основывается на механизмах привыкания (или угашения, по Павлову) ориентировочного рефлекса. Исследования показали универсальную роль ориентировочного рефлекса, с которого начинается любой вид анализа раздражений. Биологическое значение ориентировочной реакции состоит в повышении возбудимости сенсорных систем для наилучшего восприятия действующих на организм раздражений с целью установления их биологического значения. Поэтому основными факторами в возникновении ориентировочного рефлекса являются новизна, неожиданность и значимость раздражителя для организма.
В составе ориентировочной реакции выделяют два процесса: 1) начальная реакция тревоги, удивления сопровождается повышением тонуса мышц и фиксированием позы (затаивание), генерализованным изменением электрической активности разных отделов мозга (неспецифическая настройка); 2) исследовательская реакция внимания, поворот головы, глаз, ориентация рецепторов по направлению к раздражителю. Эта стадия рассматривается как период анализа раздражителя (включая условные ориентировочные поисковые реакции) и сопровождается более локальными изменениями биоэлектрической активности мозга.
Исследование динамики ориентировочного рефлекса у человека и животных показало, что он возникает не на сам стимул как таковой, а в результате сличения стимула со следом, оставленном в нервной системе предшествующими раздражителями. Если стимул и след совпадают, то ориентировочная реакция не возникает. В том случае, если информация о стимуле и хранящиеся в памяти следы от предшествующих раздражений не совпадают, то ориентировочная реакция возникает и оказывается тем интенсивнее, чем больше выражено такое рассогласование. Следовательно, конфигурация следов, оставленных в нервной системе в результате повторения каких-то раздражителей, фиксирует все их параметры. Иными словами, нервная система модификацией и комбинаторикой своих элементов строит «нервную модель» стимула (Б. Н. Соколов, 1981). Динамика построения такой нервной модели и отражается в процессах угашения ориентировочной реакции.
Отсутствие ориентировочной реакции на повторяющийся стимул свидетельствует о согласовании его нервной модели с тем реальным импульсным залпом, который направляется в головной мозг. Недавние исследования (Э. Кендел, 1980; Б. И. Котляр, 1986) установили тесную связь между сенситизацией и привыканием, которые рассматриваются в качестве двух независимых процессов. В ходе многократного предъявления раздражителей отмечено первоначальное усиление ответов (сенситизация) с последующим их подавлением (привыкание). Предположили, что оба эти процесса обеспечиваются разными нейронными сетями.
Импринтинг (запечатление). Среди форм индивидуальных адаптации особое место занимают процессы на ранних этапах постнатального развития, связанные с установлением жизненно важных контактов в гнезде, в стаде или стае, в группе или семье, в окружении родителей.
Комплекс поведенческих адаптации новорожденного, которые обеспечивают первичную связь между ним и родителями и как бы замыкают цепь преобразований эмбрионального периода, позволяя реализовать новорожденному уже сформированные механизмы восприятия и реагирования, именуют импринтингом (запечатлеванием).
К. Лоренц (1937) выдвинул оригинальную теорию импринтинга. Он считал, что молодые птицы узнают взрослых членов своего вида не инстинктивно, а с помощью импринтирования. Последнее выполняется на основе врожденной способности следовать за движущимся объектом, который попадает в их поле зрения сразу после вы-лупления.
К. Лоренц считал, что от истинного ассоциативного обучения импринтинг отличается следующими четырьмя особенностями: 1) он приурочен к ограниченному периоду жизни, именуемому «критическим или чувствительным периодом»; 2) импринтинг необратим, то есть, возникнув в критический период, он не уничтожается последующим жизненным опытом и сохраняется на всю жизнь; 3) уникальность импринтинга определяется тем, что он происходит в тот период, когда соответствующее (например, половое) поведение еще не развито. Иными словами, обучение путем импринтирования не требует подкрепления; 4) Лоренц понимал импринтинг как форму «супериндивидуального условного рефлекса», при котором запечатлеваются не индивидуальные, а видоспецифические характеристики жизненно важного объекта. Например, поведение в результате импринтирования будет направлено не к конкретной особи, которую воспринимало животное, а к целому классу организмов, к которому принадлежала импринтируемая особь.
На рис. показана установка для изучения запечатления искусственной фигуры матери.
Рис. Прибор, используемый для изучения импринтинга (реакция следования) у птиц (по А. Д, Слониму, 1976)
Движения модели утки регулируются с пульта управления внизу. Утенок следует за моделью.
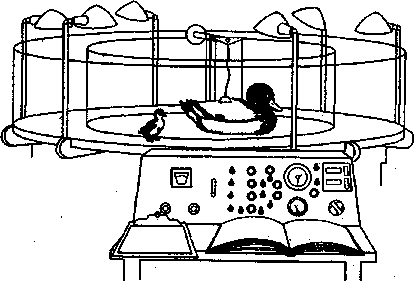
Сохранение приобретенного опыта проверяется по реакции следования утенка за муляжом взрослой утки. Такая форма обучения получила название «запечатление привязанности». Что же касается слуховых раздражений, то предполагают, что их импринтирование может иметь место еще раньше, то есть до рождения или вылупления (А, Д. Сло-ним, 1976). Свойством импринтирования обладают многие животные и насекомые, а также новорожденные дети. Причем для выработки предпочтения оказывается не -существенным длительность экспозиции объекта. Значит, связи, возникающие при запечатлевании, шире, чем. реакции следования, которую изучал К. Лоренц. Отсюда становится понятным запоминание животными местности, положения норы, гнезда и других жизненно важных ориентиров.
До сих пор спорным остается вопрос о критическом периоде импринтирования, его продолжительности и факторах, его определяющих. Расширение спектра воздействующих на организм раздражений, возрастание вероятностного характера возникновения той или иной жизненной ситуации увеличивает уровень тревожности организма и побуждает его перейти от облигатных форм обучения к факультативным.
Совершенно не разработанным оказывается вопрос о возможности импринтирования матерью своих детенышей. Например, козы, если их лишить своих детенышей только на 15 мин после родов, принимают и допускают их к себе. При удлинении этого времени до 3,5 ч козы отвергают детенышей. Такая же привязанность отмечена и у овец. Несомненно, что одной из основных функций импринтинга является установление контакта молодых особей с родителями и сородичами, то есть установление социальных отношений между молодыми и другими представителями вида. Этот период первичной социализации у незрелорождающихся животных накладывает отпечаток на всю последующую жизнедеятельность.
В самостоятельную категорию явлений запечатле-вания К. Лоренц отнес «половое импринтирование». Суть состоит в том, что самец птицы, воспитанной среди особей другого вида, став взрослым, предпочитает в

И. С. Бериташвили
качестве половых партнеров только самок этого вида, но не своего собственного. Усыновленные самцы игнорируют самку собственного вида и ухаживают за самкой, принадлежащей виду его приемных родителей. Тот факт, что импринтинг возникает задолго до созревания соответствующего поведения, подтверждается следующим наблюдением. Проигрывание песни молодым птицам влияет на песню, которую они будут петь спустя месяцы, когда достигнут половозрелости.
Это и подобного рода наблюдения — очевидное свидетельство того, что импринтинг может служить примером долговременной образной памяти (по И. С. Бериташвили), возникшей без биологического подкрепления после одноразового воздействия раздражителя. В проявлениях импринтинга используется взаимодействие индивидуального опыта и врожденных свойств молодого организма к быстрой фиксации его в механизмах памяти. Нейробиологические механизмы импринтирования, как одной из форм памяти, еще только начинают исследовать (Г. Хорн, 1988).
Подражание. Уже рассматривая примеры импринтинга, можно было обратить внимание на большую роль в формировании реакции следования выводковых птиц явлений подражания. В результате имитации (подражания) животное выполняет видотипичные действия путем непосредственного наблюдения за действиями других животных своего вида. Эти последние являются побуждающим фактором для одновременного ухода всех членов стада или стаи от хищника, для собирания пищи, отдыха и т. д. Путем имитационного научения животные на основе общения формируют поведение, которое вполне укладывается в рамки видового стереотипа.

Л. А. Орбели
Особенно это характерно для молодых животных, которые путем имитации родительского поведения обучаются различным проявлениям поведенческого репертуара своего вида. Такое имитационное научение названо облигатным. Л. А. Орбели (1949) считал имитационное поведение главным охранителем вида, ибо громадное преимущество заключается в том, что «зрители, присутствующие при акте повреждения члена их же стада или сообщества, вырабатывают рефлекторные защитные акты и таким образом могут в будущем избежать опасности. Таким образом, облигатное, неассоциативное обучение обеспечивает жизнедеятельность особи на первых этапах ее самостоятельною существования, а также закладывает основы видоспецифического характера поведения. Можно сказать, что генетическая программа с помощью видоспецифического обучения создает основы для сохранения целостности популяции.
АССОЦИАТИВНОЕ ОБУЧЕНИЕ
Классические условные рефлексы. Как уже отмечалось, формы ассоциативного обучения характеризуются совпадением во времени (ассоциацией) какого-либо воспринимаемого индифферентного раздражителя — внешнего или внутреннего — с деятельностью самого живого организма. Биологический смысл такой ассоциации — условного рефлекса — в его сигналъности, то есть в приобретении этим раздражителем роли предупреждающего фактора, сигнализирующего наступление предстоящих событий и подготавливающего организм к взаимодействию с ними. И. П. Павлов разработал учение об условных рефлексах в основном на примере рефлекторного слюноотделения. Индифферентный раздражитель сочетался с безусловным агентом, вызывающим слюноотделительный рефлекс. В этом случае возникала такая цепь событий: какой-то посторонний раздражитель — безусловное раздражение ротовой полости при попадании в нее пищи — безусловное слюноотделение. Попадание пищи в ротовую полость именовалось подкреплением. По мере сочетаний этих агентов первый раздражитель приобретал свойство условного сигнала, способного вызывать реакцию слюноотделения. Таким образом, условный сигнал как бы имитировал безусловно-рефлекторную реакцию, лишь сдвинутую во времени.
Такой принцип выработки условных рефлексов был воспроизведен и в другой экспериментальной ситуации, а именно в случае применения электро-оборонительных безусловных рефлексов. Какой-либо посторонний раздражитель, например, сочетался со слабым электро-болевым стимулом, приводящим к отдергиванию конечности у собаки. Спустя некоторое время отдергивание конечности возникало уже после предъявления условного сигнала, то есть опять же имело место сигнальное воспроизведение безусловного рефлекса. Конечно же, полного тождества безусловной реакции по условному сигналу не возникает. Правильнее говорить в этом случае об их подобии, а не об идентичности. Такие условные рефлексы еще называют условными рефлексами первого типа (Ю. М. Конорски, 1970). Они получили наименование классических. Благодаря этим условным рефлексам обеспечивается первичная ориентация животного по признакам окружающей среды еще с самого начала любого поведенческого акта. С помощью классических условных рефлексов животное адаптируется к внешней среде. Принимая во внимание модулирующее значение физиологических потребностей, в этих случаях животное все же выступает в качестве достаточно пассивного участника событий, не имеющего возможность кардинально изменить их последовательность. Деятельное же, активное начало сведено к достаточно простым безусловнорефлекторным актам.
Инструментальные условные рефлексы. Некоторые ученые в самостоятельную форму ассоциативного обучения объединяют условные рефлексы, которые строятся на основе активной целенаправленной деятельности животного. Последовательность событий в данном случае л а висит не только от внешней сигнализации, но и от поведения самого животного. Например, крыса в процессе ориентировочно-исследовательского поискового поведения и экспериментальной камере случайно нажимает лапой на педаль, за что вознаграждается пищей. По мере таких совпадений вырабатывается прочная связь: нажатие на педаль — пища. Затем экспериментатор перед очередным «произвольным» нажатием на педаль включает какой-либо посторонний раздражитель. После нескольких сочетаний всей этой цепи раздражений устанавливается связь: сигнал — нажатие на педаль — пища. Активное взаимодействие животного со средой достигается благодаря включению в эту цепь среднего звена — нажатия животным на педаль, ибо от своевременности и качества выполнения данной реакции зависит успешность поведенческого, акта в форме достижения пищевого подкрепления. При этом следует иметь в виду, что в отличие от классических условных рефлексов сам по себе двигательный акт не имеет генетической связи с получением пищи. Эта связь формируется в процессе предварительной тренировки.
Впервые инструментальные условные рефлексы стали изучаться в павловской лаборатории Ю. М. Конорски и Я. Миллером (1936), которые первоначально дали, им название — условные рефлексы второго типа. Затем их стали называть инструментальными условными рефлексами, оперантным поведением. По существу этот вид обучения не отличается от обучения с помощью проб и ошибок. Сущность инструментальной деятельности заключается в изменении взаимоотношений организма со средой, что происходит либо при изменении его положения в пространстве (локомоторная деятельность), либо при воздействии организма на окружающие предметы (манипуляторная деятельность).
Одним из распространенных примеров такого обучения является выработка у животных навыков ориентации в лабиринте. Животное помещается в лабиринт (рис. 12), в котором находится пищевая приманка. После неоднократного прохождения лабиринта путем его тщательного исследования животное обучается кратчайшим путем достигать местоположения приманки. Кривая обучения обычно строится на основе скорости достижения приманки и/или количества допущенных ошибок. Лабиринтные навыки довольно быстро становятся автоматизированными и стереотипными.
Было отмечено, если крысу держать в лабиринте еще до обучения, но без всякого подкрепления, то в дальнейшем она обучается быстрее и точнее. Считается, что при обследовании лабиринта животное уже приобретает оп ределенный опыт (перцептивное латентное обучение), который используется для целей нахождения кратчайшего пути в лабиринте.
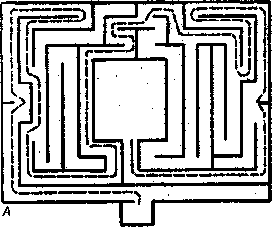
Рис. Лабиринты: А — план первого лабиринта, применявшегося в зоопсихологических исследованиях (лабиринт Смолла); Б — лабиринт из «мостиков» (по К, Э. Фабри, 1976)
Пунктиром показана траектория побежки крысы.
Другим примером является оперантное обучение в «проблемном ящике». При таком обучении животные начинают использовать разнообразные реакции, имеющиеся в их природном репертуаре, а не навязанные экспериментатором. Например, кошка, помещенная в закрытую камеру, пытается убежать, безостановочно двигается по камере и путем перебора поисковых реакций случайно поворачивает щеколду и получает доступ к пище. Таким образом, системы классических и инструментальных условных рефлексов, участвуя в сложной конструкции адаптивного поведения, значительно расширяют приспособительные возможности живого организма, который начинает выступать в качестве активного фактора взаимодействия со средой.
КОГНИТИВНОЕ ОБУЧЕНИЕ
Образное (психонервное) поведение. И. С. Бериташвили начиная с 1926 г. изучал поведение животных по разработанному им методу свободного перемещения в экспериментальном манеже. Полученные И. С. Бериташвили (1975) факты было невозможно объяснить с позиций теории условнорефлекторного обучения. Суть представлений этого ученого состоит в следующем: у высших позвоночных животных при первом же восприятии местоположения пищи создается образ или конкретное представление пищи и ее местоположения в данной среде. Этот образ сохраняется и каждый раз, когда он репродуцируется при восприятии данной среды или какого либо его компонента, животное производит такое же ориентировочное движение головы, как и при непосредственном восприятии, ведет себя точно так, как при восприятии, то есть идет к месту пищи, обнюхивает его и, если находит пищу, съедает ее. Такое психонервное поведение, направленное образами, стали называть произвольным в отличие от условнорефлекторного поведения, которое И. С. Бериташвили считал автоматизированным, непроизвольным.
И. С. Бериташвили описывает следующие характерные особенности психонервной деятельности.
1. Психонервная активность интегрирует элемент внешней среды в одно целое переживание, производящее целостный образ. Для этого достаточно, чтобы животное один раз восприняло эту среду.
2. Психонервный комплекс образа легко воспроизводится под влиянием только одного компонента внешней среды или раздражения, напоминающего эту среду.
3. Это воспроизведение может происходить спустя длительное время после начального восприятия жизненно важной ситуации. Иногда образ может удерживаться всю жизнь без повторного его воспроизведения.
4. При такой форме обучения устанавливаются временные нервные связи между нервным субстратом образа и двигательными центрами мозга. Индивидуально приобретенные реакции, направляемые образами, легко автоматизируются при их повторении.
5. Двигательная активность животных при репродукции образа зависит от условий его формирования, от давности возникновения, его жизненного значения, пространственных признаков ситуации.
6. Психонервная активность высших позвоночных животных преобладает над другими формами обучения и является определяющим фактором поведения.
7. Сознательная деятельность человека подчиняет себе двигательную активность психонервного процесса преставления, хотя и протекает в тесном взаимодействии со всеми другими формами поведения.
И. С. Бериташвили полагал, что нервный субстрат образной психической деятельности лежит исключительно в неокортексе.
Система нейронов, продуцирующая образы жизненно важных объектов, может производить непосредственно только ориентировочную реакцию. Все остальное поведение животного опирается на разные двигательные акты прирожденного и условнорефлекторного характера в соответствии с образом и окружающей ситуацией. Значит, образ, который лежит в основе субъективного отражения внешнего мира, — это вектор предстоящего рефлекторного поведения, непосредственная активность образа реализуется через ориентировочную реакцию, которая регулирует направление поведения животного.
Согласно И. С. Бериташвили, образ фиксируется в памяти и извлекается оттуда для удовлетворения господствующей биологической потребности.
Основные отличия данной формы психонервного поведения от закономерностей условнорефлекторной деятельности состоят в следующем (И. С. Бериташвили, 1984).
1. В отличие от классических условных рефлексов, которые требуют повторяемости, психонервный образ формируется сразу после одного осуществления индивидуального поведения.
2. Условный рефлекс, не сопровождающийся безусловным подкреплением, осуществляется в течение длительного времени и гаснет постепенно. Психонервный образ, не сопровождающийся удовлетворением соответствующей биологической потребности, быстро перестает вызывать индивидуально направленное поведение.
3. Если условный рефлекс может образовываться и в отсутствие коры, то целостные представления могут возникнуть только при наличии интактной коры, и прежде коего лобных отделов коры больших полушарий.
4. Условный рефлекс подразумевает формирование к ременной связи между сенсорными и моторными элементами. Психонервное поведение обусловлено образованием новых действенных нервных кругов из сенсорных корковых нейронов.
5. Психонервная деятельность определяет включение; и выключение условных и безусловных рефлексов и последовательность их протекания в данном ряду.
Существенно, что индивидуальное поведение, первично направляемое психонервным образом, при повторной тренировке автоматизируется и осуществляется по всем закономерностям условнорефлекторного обучения.
Элементарная рассудочная деятельность животных. Поведенческие адаптации, с точки зрения Л. В. Крушинского (1977), строятся на трех основных механизмах: инстинктивном, механизме обучаемости и рассудочной деятельности (разуме). Хотя последние способности животных противопоставляются обучению, данную форму индивидуальной адаптации целесообразно рассмотреть в общей системе форм поведения особи. На рис. приведена схема взаимосвязи основных элементарных компонентов, участвующих в формировании поведенческого акта. В результате интеграции компонентов формируется целостный поведенческий акт, который можно определить как унитарную реакцию. Если на ранних этапах филогенетического развития поведение формируется под ведущим влиянием инстинктов, то по мере усложнения нервной системы большую роль начинают играть различные формы обучения. Однако дальнейшая дифференциация конечного мозга обеспечивает элементарной рассудочной деятельности все большую роль в обучении.
Рис. Схема соотношения основных элементарных компонентов поведения (по Л. В. Крушинскому, 1977). Модификация поведения возможна для каждого компонента в пределах определенной «нормы реакций».
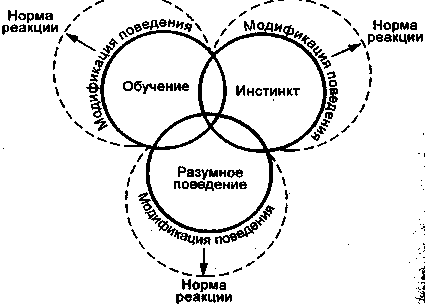
Следовательно, чем выше уровень элементарной рассудочной деятельности животных, тем пластичнее и адаптивнее любая унитарная реакция и все поведение в целом.
Наиболее характерное свойство элементарной рассудочной деятельности животных заключается в их способности улавливать простейшие эмпирические законы, связывающие предметы и явления окружающей среды, и возможность оперировать этими законами при построении программ поведения в новых ситуациях. Иными словами, важнейшая функция рассудочной деятельности состоит в отборе той информации о структурной организации среды, которая необходима для построения программы наиболее адекватного акта поведения 11 данных условиях. Автор показал, что способность к эстраполяции направления движения раздражителя может рассматриваться как одно из элементарных проявлений рассудочной деятельности животных. Основной экспериментальный подход состоит в следующем. Животное должно находить кормушку А, двигающуюся прямолинейно с постоянной скоростью. Первоначальный отрезок ее движения происходит в поле зрения животного, затем кормушка А скрывается за преградой.
Животное устремляется к месту выхода из непрозрачного коридора кормушки А с кормом. При этом животное определяет 1) из соотношения с неподвижными точками пространства изменение положения кормушки А, то есть направление ее движения, и 2) направление движения кормушки А после ее исчезновения за укрытием. Таким образом, используя свою «систему отсчета», животное экстраполирует направление смещения кормушки А с кормом, то есть использует предварительно воспринятую тактику изменений в среде для построения логики своего будущего поведения. Различные тесты, предъявлявшиеся животным разного филогенетического уровня, выявили их способность улавливать простейшие законы пространства, времени, движения.
Л. В. Крушинский считал, что чем больше размер мозга, а значит, и количество нейронов, тем большую информацию об окружающей среде он может получить. Наличие избыточного числа нейронов с разной функциональной активностью позволяет животному улавливать некоторые законы природы.
В процессе синтеза (эвристического уровня деятельности мозга) происходит сличение уловленных в данный момент законов с теми, которые были познаны в предшествующей жизни. В результате такого сравнения осуществляется выбор наиболее адекватного пути решения задачи. Совпадение новой информации с записанной в долговременной памяти облегчает решение задачи.
Исходя из особенностей поведения животных в сообществах, автор приходит к выводу, что степень многообразия и пластичности отношений в сообществе животных находится в зависимости от уровня развития их рассудочной деятельности, выполняющей особую роль в эволюции популяций.
Вероятностное прогнозирование. Под вероятностным прогнозированием понимается предвосхищение будущего, основанное на вероятностной структуре прошлого опыта и информации о наличной ситуации. Оба эти фактора служат основой для создания гипотез о предстоящем будущем, причем каждой из них придается определенная вероятность. В соответствии с таким прогнозом осуществляется подготовка к действиям в предстоящей ситуации, приводящим к наибольшей вероятности достижения цели (Н. А. Бернштейн, 1966).
Способность к вероятностному прогнозированию — результат биологической эволюции в вероятностно организованной среде. Прогнозы живого организма призваны оптимизировать результаты его действия.
В естественных условиях организм сталкивается с множеством различных случайных раздражителей, которые нужно статистически обрабатывать, чтобы выявить повторяемость, то есть регулярность событий, на основании которой и формируется приспособительное поведение. Теория вероятностного обучения исходит из представлений о предсказании статистических закономерностей и выбора оптимальных стратегий поведения при обучении субъекта независимым вероятностям стохастично возникающих раздражений.
П. В. Симонов (1986) считает, что степень информированности субъекта зависит от текущей субъективной вероятности достижения результата с учетом наличных средств и оценки проблемной ситуации. В связи с этим из реальной окружающей среды выделяют и ту актуальную среду организма, от которой непосредственно зависит удовлетворение потребностей и достижение целей.
Высшие позвоночные и человек в своей деятельности непрерывно опираются на вероятностное прогнозирование. Когда человек переходит улицу, он прогнозирует ситуацию на проезжей части и интервал времени для безопасного пересечения дороги. А водитель транспорта в это время прогнозирует поведение пешеходов, смену сигналов светофора и свои собственные действия по управлению автомобилем.
Животные прогнозируют вероятность поиска пищевого объекта, одновременно прогнозируя поведение хищника, чтобы не стать жертвой собственной «беспечности».
В целом вероятностное прогнозирование деятельности может иметь разные формы (И. М. Фегенберг, Г. Е. Журавлев, 1977):
1. Прогнозирование разных форм независимых от субъекта событий.
2. Прогнозирование своих ответных активных действий.
3. Прогнозирование целенаправленных действий не только в соответствии с их частотой в прошлом опыте, но и с их актуальной значимостью и предполагаемым результатом.
4. Поведение в общении с другими организмами предполагает использование гипотез о наиболее вероятных действиях своих активных партнеров.
5. Прогнозирование действий и целей с учетом собственных энергетических затрат.
Таким образом, когнитивные формы обучения, опираясь на весь широкий репертуар более простых форм облигатного и ассоциативного обучения, выступают в качестве одного из фундаментальных механизмов высшей нервной деятельности животных и человека.