

МОЗ УКРАЇНИ
КОВЕЛЬСЬКИЙ МЕДИЧНИЙ КОЛЕДЖ

МЕТОДИЧНА РОЗРОБКА ПРАКТИЧНОГО ЗАНЯТТЯ №4
ПО ТЕМІ:
«Вегетативні органи рослин. Пагін, листок як органи рослини»
Підготувала викладач
Совтус І.М.
Практичне заняття №4
Тема: «Вегетативні органи рослин. Пагін, листок як органи рослини»
Знати:
Вегетативні органи рослин: пагони, листки, їх морфологія та метаморфози.
Морфологічне описування вегетативних органів рослин.
Практичні навички:
проведення морфологічного вивчення вегетативних органів на живих рослинах, гербарних та сухих зразках;
розпізнавання метаморфоз вегетативних органів;
морфологічне описування пагона (стебла, листків);
замальовування об’єкта, що вивчається.
СТЕБЛО
Стебло вивчається як частина пагона. Стебло – вісь пагона, що складається з вузлів та міжвузлів і несе листки та бруньки. Наростання стебла відбувається за рахунок верхівкової, латеральних та вставної меристем.
Стебло має більш-менш правильну циліндричну форму і радіальну симетрію в розміщенні тканин, хоча на поперечному зрізі воно може бути округлим, сплющеним і крилатим, три-, чотири- та багатогранним тощо.
Функції стебла:
провідна (нагору від коренів до листків, квіток і плодів проводить воду з мінеральними речовинами, униз від листків – розчин органічних речовин);
осьова (розташування листків, квіток, плодів);
запасна (м’ясисті стебла більшості рослин запасають воду й органічні речовини: бульби картоплі, цибулини цибулі, часнику, стебла кактусів, цукрової тростини);
вегетативне розмноження (бульбами, кореневищами, вусами, відводками, черешками, цибулинами тощо).
За розташуванням стебел у просторі розрізняють:
ортотропні – прямостійні стебла (соняшник, кукурудза, пшениця);
плагіотропні – повзучі (вкорінюються у вузлах: суниця, конюшина повзуча, жовтець повзучий) та сланкі (не вкорінюються: огірок);
виткі (іпомея, хміль);
чіпкі (мишачий горошок, горох посівний, виноград).
Стебла можуть бути:
трав’янистими – більше живих тонкостінних клітин, napeнхімних тканин;
здерев’янілими – більше мертвих клітин із товстими клітинними стінками, що виконують провідну і механічну функції.
Анатомічна будова стебла відповідає його основним функціям, тобто воно має розвинену систему:
провідних тканин – зв’язує всі системи та органи рослини і забезпечує транспорт води і поживних речовин;
механічних тканин – забезпечує виконання стеблом його осьової та опірної функцій.
Структура стебла як осьової частини рослинного організму формується переважно внаслідок діяльності його верхівкової (апікальної) меристеми (мал. 19, 1) або ініціальних клітин конуса наростання (точки росту). Конус наростання – це нерозгалужена верхівка стебла до 1,5-2 мм завдовжки. Клітини його щільно зімкнені, протопласт без вакуолей, з великим ядром і густим вмістом. Особливістю клітин є активна здатність до поділу.
Таким чином, утворюється велика кількість клітин, наростає маса стебла. Морфологічно клітини конуса наростання однорідні, але генетично вони різні і дають початок постійним тканинам центрального циліндра первинної кори і епідерми.
Про походження і розвиток тканин з окремих гістогенів розроблені численні теорії, але найбільшої популярності набули теорія гістогенів Ганштейна і теорія туніки і корпуса Шмідта.
1. За теорією Ганштейна, у конусі наростання кореня є три зони ініціальних клітин, які шляхом поділу утворюють гістогенні зони: дерматоген, периблему і плером, з яких відповідно виникають епіблема, первинна кора і центральний циліндр. Ця теорія більше підходить для кореня, а для стебла не цілком прийнятна, оскільки не завжди чітко розмежовуються зони ініціальних груп клітин, з яких формуються плером і периблема, або тканини, що утворюються з них, не відповідають накресленій схемі. Наприклад, мезофіл листка пшениці утворюється не з периблеми, а з дерматогену.
2. Тому розвиток стебла краще пояснюється теорією туніки і корпуса. За теорією Шмідта, у конусі наростання є дві зони ініціальних клітин, що під час поділу утворюють два гістогени: туніку і корпус. Туніка має один або кілька шарів. Клітини її поділяються перпендикулярно до поверхні стебла і утворюють епідерму, а в деяких рослин формується кілька зовнішніх шарів первинної кори. Усю внутрішню частину конуса наростання складає корпус. Його клітини діляться в різних напрямках і розміщуються не пошарово, а безладно. Корпус дає початок розвитку постійним шарам первинної кори і центрального циліндра.
Нижче конуса наростання клітини подовжуються, збільшуються в розмірах і набувають певної постійної форми. Саме завдяки цій частині відбувається найінтенсивніший ріст, тому її називають зоною росту. Тут помітний поділ периферійних клітин, що утворюють горбочки зародкових листків (примордії, мал. 19, 3, 4). Клітини зародкових листків більші за розмірами від клітин апікальної меристеми (мал. 19, 2), мають сформовану вакуолю.
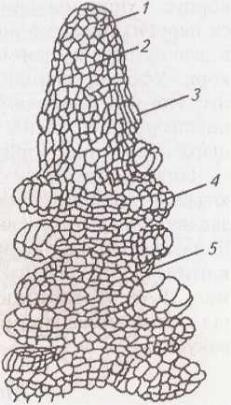
1
–
апекс;
2
–
клітини апікальної меристеми;
3-4
–
листкові зачатки;
5
–
зародкові міжвузляМал. 19. Верхівка пагона в повздовжньому розрізі:
Первинна будова стебла однодольних рослин
Первинна будова стебла однодольних рослин зберігається все життя. Розвиток її починається з функціонування первинних меристем – туніки, корпуса і прокамбію. Всі тканини, що утворюються з них і складають стебло первинної будови, за походженням є первинними. Первинна будова стебла різноманітна і залежить від розвитку в певному середовищі виду рослини, умов вирощування тощо. Але завжди в ній можна виділити три основні частини стебла:
епідерму,
первинну кору,
центральний циліндр.
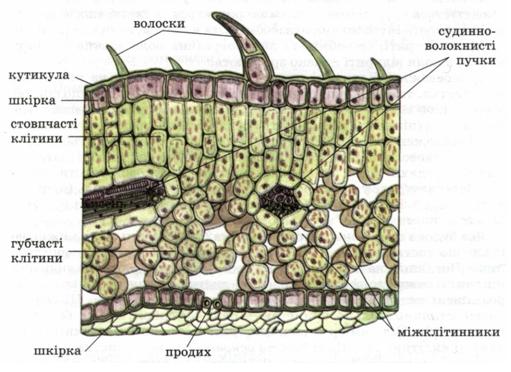
Епідерма стебла (мал. 20, 1) не має звивистих клітинних оболонок, містить менше продихів і має тонший кутикулярний шар, ніж у листків. Під епідермою суцільним шаром (соняшник) або по кутах (шавлія) окремими ділянками залягає кутова, або пластинчаста, коленхіма.
Первинна кора в багатьох рослин займає значну частину стебла і складається з коленхіми (мал. 20, 2), корової паренхіми (мал. 20, 3) і ендодерми (мал. 20, 4). Вона зміцнює молоде стебло. Її клітини діляться і розтягуються разом з іншими тканинами стебла.
Корова паренхіма – це жива тканина, її клітини з тонкими оболонками, заповнені протопластом. У клітинах зовнішніх шарів містяться хлоропласти, які виконують функцію фотосинтезу. Зберігаючи свою життєвість, вони можуть утворювати фелоген. Клітини внутрішніх шарів первинної кори позбавлені хлоропластів і часто є місцем запасання поживних речовин.
Стебла водних рослин мають добре розвинену аеренхіму – паренхіму кори з великими міжклітинниками. У паренхімі кори можуть формуватися смоляні ходи, молочники, вмістилища видільних тканин і кам’янисті клітини, відкладаються кристали кальцію оксалату.
Внутрішній шар клітин первинної кори утворює ендодерма.
Клітини її щільно зімкнені, у деяких рослин радіальні стінки дерев’яніють. У сучасних рослин вона краще розвинена в злаків та інших однодольних рослин, особливо водних. Для багатьох покритонасінних рослин ендодерма не характерна. Замість неї розвивається крохмалоносна піхва, клітини якої заповнені крохмальними зернами, що не використовуються в процесі життєдіяльності органа, а відіграють роль статолітів (це вид амілопластів – лейкопласти, в яких відбувається синтез крохмалю, переважають у запасаючих тканинах різних органів рослин, особливо в бульбах, цибулинах, кореневищах – нагромаджують в них вторинний (запасний) крохмаль)
С
1
— епідерма;
2
—
коленхіма;
3
—
корова паренхіма;
4
— ендодерма;
5
— перицикл;
6
—
флоема;
7
— прокамбій;
8
— ксилема;
9
— паренхіма центрального циліндра
Мал. 20. Первинна будова стебла дводольної рослини:
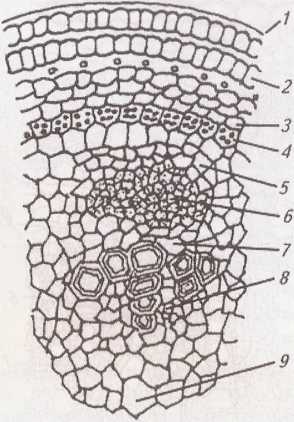
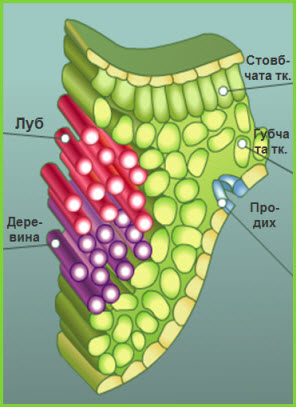
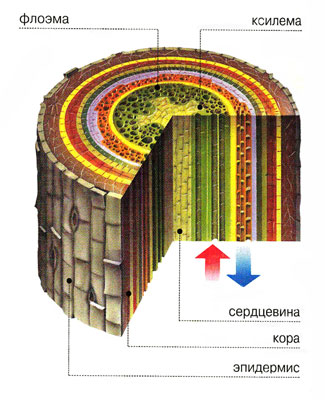
Серцевина з’єднується із первинною корою за допомогою паренхіми серцевинних променів. Завдяки їм відбувається надходження поживних речовин і газообмін у горизонтальному напрямку. У серцевину занурені провідні пучки, які розвиваються з прокамбіальних тяжів, або суцільні циліндри флоеми (мал. 20, 6) і ксилеми (мал. 20, 8), розділені камбієм, що утворився із суцільного прокамбіального циліндра (мал. 20, 7). Отже, залежно від характеру закладання прокамбію (у вигляді окремих тяжів чи суцільного кільця) виникає безпучковий (мал. 21) або пучковий (мал. 22) тип будови стебла.
Мал.
21.
Безпучковий
тип
будови стебла
льону:
1
—
епідерма;
2
— паренхіма кори;
3
— первинні луб’яні волокна;
4
— вторинна флоема;
5
— камбій;
6
— вторинна ксилема;
7
— первинна ксилема
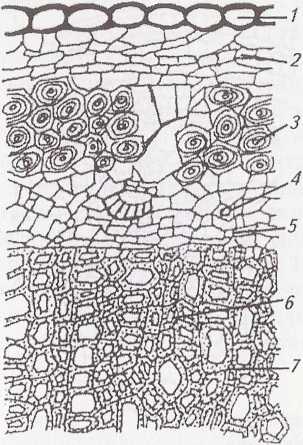
Man.
22.
Пучковий
тип будови стебла хвилівника:
1
— кутикула;
2
— епідерма;
3
—
коленхіма;
4
— паренхіма кори;
5
— ендодерма;
6
— склеренхіма;
7
— провідний пучок;
8
—
флоема;
9—
міжпучковий камбій;
10—
пучковий камбій;
11
— ксилема;
12
— серцевинний промінь;
13
—
серцевина
У процесі поділу клітин прокамбію диференціюються елементи первинної ксилеми і флоеми: трахеїди, судини, ситоподібні трубки, клітини-супутниці. Ксилема складається з протоксилеми і метаксилеми, а флоема – з протофлоеми і метафлоеми. Першими виникають спіральні і кільчасті судини, пізніше складніші елементи: драбинчасті і сітчасті судини, ситоподібні трубки з клітинами-супутницями. Прокамбій в однодольних рослин повністю витрачається на формування ксилеми і флоеми. Так, виникають закриті провідні пучки. Якщо прокамбій зберігається і дає початок пучковому камбію, який відтворює ксилему до центру, а флоему – до периферії, то такий пучок називається відкритим. Вони властиві дводольним рослинам.
Особливості анатомічної будови стебла однодольних. Однодольні рослини представлені трав’янистими й деревними формами, однорічними й багаторічними видами. Вони мають різноманітну анатомічну будову стебла.
У рослин з верхівковим ростом і виповненим стеблом (півникових, лілійних) чітко виражені епідерма, первинна кора і центральний циліндр. Епідерма (мал. 23, 1) має подовжені клітини з продихами. Коленхіма, як правило, відсутня, іноді є виражена склеренхіма (мал. 23, 2). Далі залягає хлорофілоносна паренхіма первинної кори, яка бере участь у фотосинтезі. Внутрішній шар первинної кори утворює ендодерма (крохмалоносна піхва). Глибше розміщена склеренхіма, що виникає з клітин перициклу. Основну частину стебла становить серцевина, в якій безладно розміщені провідні пучки закритого колатерального типу (мал. 23, 4). Особливу будову мають стебла злаків, наприклад кукурудзи. Зверху стебло вкриває один шар епідерми з продихами. Первинна кора інколи виражена тільки вузькими прошарками хлорофілоносної паренхіми. Нерідко склеренхіма оточує ці ділянки, і годі вона знаходиться безпосередньо під епідермою.
Склеренхімне кільце має перициклічне походження і складає зовнішній шар центрального циліндра. Периферійне розміщення склеренхімного кільця надає міцності високому стеблу. Найбільшу частину стебла заповнює основна паренхіма, пронизана провідними пучками (мал. 23, Б).
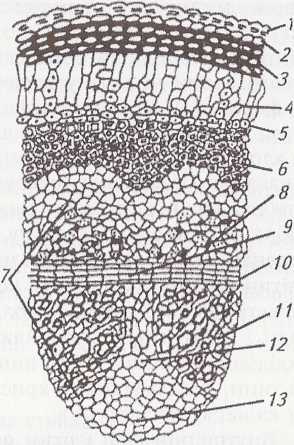
Мал.
23.
Анатомічна
будова (поперечний зріз):
А
— стебла
кукурудзи:
1
—
епідерма;
2
— склеренхіма;
3
— основна паренхіма;
4
— провідний пучок закритого типу;
Б
—
провідного пучка:
5
— тонкостінна паренхіма;
6
— склеренхімна піхва;
7
— флоема;
8
— пориста судина;
9
– кільчасто-спіральна судина;
10
—
ксилемна паренхіма;
11
— порожнина розриву;
12
— ксилема
Провідні пучки утворюються з прокамбіальних тяжів. Доки функціонує прокамбій і відчленовуються елементи флоеми, ксилеми (мал. 23, 7, 12), механічних і паренхімних тканин, стебло активно потовщується. З часом прокамбій повністю втрачає меристематичні властивості і потовщення стебла уповільнюється чи припиняється. Флоема складається із ситоподібних трубок, клітин-супутниць і флоемної паренхіми. Вона орієнтується до периферії стебла. Ксилема складається з двох великих пористих судин, розмежованих товстостінною паренхімою, до якої прилягають кільчасто-спіральна і кільчаста судини. Під ними є порожнина розриву (мал. 23, 11), а з боків – ксилемна паренхіма (мал. 23, 10). Ксилема від центру підковою охоплює флоему і разом вони утворюють колатеральний провідний пучок. Його оточує склеренхімна піхва (мал. 23, 6).
Провідні пучки в однодольних розміщені безладно. Більші за розмірами знаходяться ближче до центру, а менші – до периферії; їх тут більше із потужнішою склеренхімною піхвою.
Рослини з інтеркалярним ростом і порожнистим стеблом (більшість злаків – пшениця, жито, ячмінь – мал. 24) мають певні особливості анатомічної будови. Зверху стебло вкрите епідермою, клітинні оболонки якого дерев’яніють під кінець вегетації. Первинна кора представлена невеликими ділянками хлорофілоносної паренхіми (мал. 24, 4) між провідними пучками. Коленхіма і крохмаленосна піхва відсутні, добре розвинена склеренхіма. Найбільша частина соломини виповнена основною паренхімою. З розвитком стебла руйнується її центральна частина, а утворена порожнина відтісняє решту паренхіми до периферії. Провідні пучки розміщуються через це у двох колах.
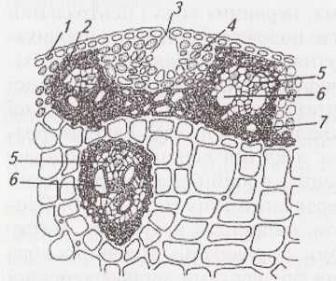
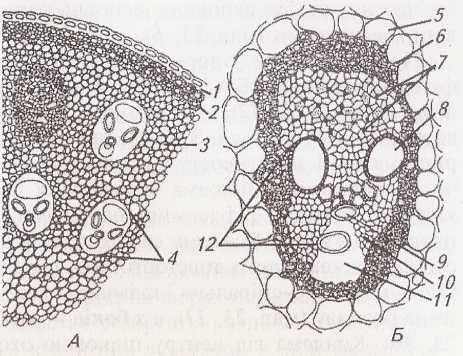
Мал.
24.
Будова
соломини жита:
1
–
епідерма;
2
–
склеренхіма;
3
–
продих;
4
–
хлорофілоносна паренхіма;
5
–
флоема;
6
–
ксилема;
7
–
паренхіма
центрального циліндра
Внутрішні пучки дещо більші, зі слаборозвиненою склеренхімою, зовнішні менші за розміром і оточені потужною склеренхімною піхвою. Зовнішній шар центрального циліндра представлений перициклом, з якого утворилося склеренхімне кільце. Склеренхіма, перекидаючись від одного пучка до іншого і зливаючись зі склеренхімними піхвами та радіальними відгалуженнями, утворює суцільний каркас механічної тканини, яка надає стеблу міцності і протистоїть згинанню.
Деякі однодольні рослини (драцена, юка, алое, агава) характеризуються вторинним ростом, але він відбувається не за рахунок камбію, а внаслідок виникнення паренхімної тканини. Закладається кільце стовщення у внутрішніх шарах паренхіми кори.
Наслідком його діяльності є утворення незначної частини паренхіми і основної маси вторинної деревини, в якій формуються концентричні (амфівазальні) провідні пучки. У таких пучках ксилема представлена трахеїдами.