

Методика окраски
Мазки, фиксированные в метиловом спирте, окрашивают раствором (1 мл готовой жидкой краски + 2 мл основного буферного раствора + 47 мл дистиллированной воды) в течение 40—120 мин (продолжительность окрашивания подбирают эмпирически). Пользуются фосфатным буфером, но рН буфера зависит от вида мазка: для мазка костного мозга — 5,8 — 6,0, для мазка крови — 6,4 — 6,5, для выявления простейших — 6,8, малярийного плазмодия — 7,0 — 7,2.
Ополаскивают в дистиллированной воде, высушивают и исследуют при иммерсии.
Результат окраски
При окрашивании простейших их цитоплазма приобретает голубой цвет, а ядра — красно-фиолетовый.
20. Грибы относятся к царству Fungi (Mycetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофильные) эукариотические микроорганизмы с клеточной стенкой.
Грибы имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную, ригидную клеточную стенку, состоящую из нескольких типов полисахаридов, а также белка, липидов и др.
Некоторые грибы образуют капсулу.
Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы. Грибы являются
грамположительными микробами, вегетативные клетки - не-кислотоустойчивые.
Грибы состоят из длинных тонких нитей (гиф), сплетающихся в грибницу, или мицелий. Гифы низших грибов - фикомицетов - не имеют перегородок. У высших грибов - эумицетов - гифы разделены перегородками; их
мицелий многоклеточный.
Различают гифальные и дрожжевые формы грибов. Гифальные (плесневые) грибы образуют ветвящиеся тонкие нити (гифы), сплетающиеся в грибницу, или мицелий (плесень). Гифы, врастающие в питательный субстрат, называются вегетативными гифами (отвечают за питание гриба), а растущие над поверхностью субстрата - воздушными или репродуктивными гифами (отвечают за бесполое размножение) Гифы низших грибов не имеют перегородок. Они представлены многоядерными клетками и называются ценоцитными.
Гифы высших грибов разделены перегородками, или септами с отверстиями. Дрожжевые грибы (дрожжи), в основном, имеют вид отдельных овальных клеток (одноклеточные грибы). По типу полового размножения они распределены среди высших грибов - аскомицет и базидиомицет. При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий) в виде цепочек удлиненных клеток - "сарделек". Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом - почкованием или делением. Грибы размножаются спорами половым и бесполым способами, а также вегетативным путем (почкование или фрагментация гиф). Грибы,
размножающиеся половым и бесполым путем, относятся к совершенным. Несовершенными называют грибы, у которых отсутствует или еще не описан половой путь размножения. Бесполое размножение осуществляется у грибов с помощью эндогенных спор, созревающих внутри круглой структуры - спорангия, и экзогенных спор - конидий, формирующихся на кончиках плодоносящих гиф.
Среди грибов, имеющих медицинское значение, выделяют 3 типа: зигомицеты (Zygomycota), acкомицеты (Ascomycota) и базидиомицеты (Basidiomycota). Зигомицеты: низшие грибы, вызывают зигомикоз легких, головного мозга.
Аскомицеты: высшие совершенные грибы, к ним относится возбудитель эрготизма, Микозы, дерматомикозы, пневмония, кератоз, аспергиллез. Дейтеромицеты: несовершенные грибы, род Candida, поражающие слизистые оболочки и внутренние органы. Заболевания:
1. Поверхностные микозы (кератомикозы) — поражения поверхностных слоев кожи и волос:Malassezia furfur, Cladosporium.
2.Эпидермофитии (эпидермомикозы) — поражения эпидермиса, кожи и волос:Epidermophyton floccosum,Microsporum canis.
3.Подкожные, вовлекающие в процесс дерму, подкожные ткани, мышцы: Sportrichum schenckii.
4.Системные, или глубокие, микозы, при которых поражаются внутренние органы и ткани: Coccidioides immitis, Histoplasma capsulatum/
5. Оппортунистические микозы: Candida, Aspergillus, Mucor.
6. Аллергии, вызванные грибами (пневмоаллергии и дермоаллергии).
7. Микотоксикозы — пищевые интоксикации, вызванные токсинами грибов.Лабораторная диагностика: микроскопическое, микологическое, аллергическое, серологическое исследования. Материал для исследования: гной, мокрота, волосы, кожа, кровь. Микроскопическое: микроскопия нативных препаратов: изучение строения гриба, расположение спор, культуральная принадлежность. Для окраски – методы Грамма, Романовского, Циля-Нильсена. Культуральное (микологическое) исследование: выделение чистой культуры, плотные, жидкие пит. среды.
21. Актиномикоз — хроническое заболевание, вызываемое различными видами актиномицетов. Впервые описано у человека Йзраели в 1879 г., а друзы актиномицета обнаружены Лангенбеком в 1845 г.
Морфология и биологические свойства. Возбудителями актиномикоза являются различные виды лучистых грибов: Actinomyces israeli, A. bovis, A. albus, A. violaceus. Актиномицеты имеют одноклеточный, тонкий, ветвящийся мицелий, который быстро распадается на палочковидные клетки различной длины. Встречаются короткие палочки и кокки, колбовидные и шаровидные вздутые клетки. Спор не образуют, грамположительны. У человека в выделениях из пораженных тканей обнаруживаются друзы, центр которых состоит из войлокообразного скопления ветвящихся нитей мицелия, окруженного лучами с колбообразно вздутыми концами. На плотных питательных средах актиномицеты образуют гладкие или слегка бугристые колонии тестообразной консистенции, бесцветные. Растут при 35—37°С, условные анаэробы. Устойчивость. Во внешней среде устойчивы, выдерживают нагревание до 60°С в течение часа. Патогенез и клиника. Заражение происходит через поврежденную кожу или слизистые оболочки при уколе соломой, шипами цветов, укусе лесного клеща (экзогенная инфекция). Наиболее частый путь заражения эндогенный, так как актиномицеты обычно обитают на слизистой оболочке рта, верхних дыхательных путей и желудочно-кишечного тракта. При травмах, воспалительных процессах полости рта и десен они могут вызвать заболевание. На месте проникновения актиномииетов возникают плотные, деревянистой консистенции, малоболезненные инфильтраты — узлы, кожа над которыми вначале бывает синюшной, а затем багровой. Инфильтраты размягчаются, нагнаиваются и вскрываются с образованием долго не заживающих свищей. После заживления остаются плотные втянутые рубцы. Обычно поражаются мягкие ткани головы, шеи, языка, а также органы грудной клетки, брюшной полости, малого таза и половые органы. Течение заболевания тяжелое, особенно при поражении внутренних органов, и часто осложняется вторичной гнойной инфекцией. Иммунитет. После перенесенного заболевания нестойкий. Микробиологическая диагностика. Микроскопическому исследованию подвергают гной из свищей, мокроту, мочу, пунктаты инфильтратов, кусочки органов из трупов. Из исследуемого материала выбирают желтоватые комочки, помещают на предметное стекло в каплю 10— 20% щелочи, слегка подогревают и микроскопируют. Для актиномикоза характерно наличие каменисто-плотных комочков, хрустящих, как песок, при надавливании и содержащих внутри друзы. Для выявления анаэробных актиномицетов посев исследуемого материала производят в высокий столбик 1—2% сахарного агара, который заливают вазелиновым маслом и выращивают при 37°С. Аэробные актиномицеты культивируют на чашках с агаром. Антитела в сыворотке крови больного определяют в РСК, в качестве антигена используя поливалентный актинолизат. РСК положительна в 80—88% случаев актиномикоза. Внутрикожные аллергические пробы ставят с актинолизатом, результаты учитывают через сутки. У больных с тяжелыми формами и при пониженной реактивности они могут быть отрицательными. Профилактика и лечение. Предупреждение актиномикоза связано с санацией и гигиеной полости рта, лечением кариозных зубов, пиореи и различных воспалительных процессов слизистой оболочки полости рта и миндалин. Важна также профилактика травм кожи, особенно стоп у жителей сельской местности. Наилучшие результаты лечения получены при комбинированном применении актинолизата и антибиотиков: пенициллина, стрептомицина, тетрациклинов. Используют витаминотерапию и переливание крови.
22. Спирохеты (от лат. spira — изгиб, chatie — грива) — спирально извитые одноклеточные организмы. Число витков у спирохет может достигать 10—15
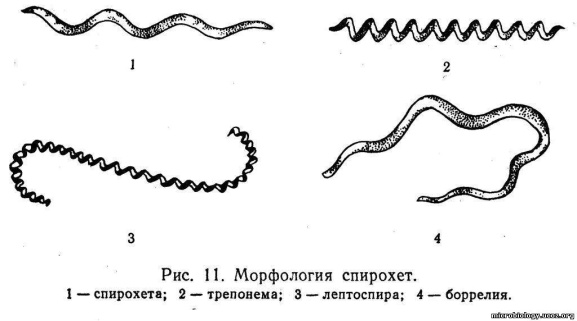
Величина их значительно варьирует: диаметр от 0,25 до 6 мкм, а длина от 7 до 500 мкм. Исследования в электронном микроскопе показали, что структура спирохет значительно сложнее, чем у бактерий. Они имеют клеточные оболочки, цитоплазматическую спираль (цитоплазматический цилиндр) и осевую нить. Внутри цитоплазмы расположены нуклеоид, образования типа мезосомы и различные гранулы. Клеточные оболочки спирохеты представляют собой комплекс из двух образований: наружной оболочки, очень тонкой, эластичной и гибкой (покров) и лежащей под ней клеточной стенки цитоплазматического цилиндра. В поверхностном слое клеточной стенки цитоплазматического цилиндра обнаружены гликопептиды. Осевая нить, вокруг которой изогнуто тело спирохет, состоит из одной, нескольких или пучка слившихся фибрилл.
В фибриллах найдено хитиноподобное вещество — кутин, которое обычно встречается только у животных. Спирохеты очень подвижны. Они могут сгибаться, сокращаться, совершать быстрые вращательные и прямолинейные движения за счет сокращений их фибриллярного аппарата. Размножаются спирохеты поперечным делением на две равноценные особи. Среди спирохет имеются возбудители инфекционных заболеваний человека: возвратного,тифа, сифилиса и лептоспирозов. Некоторые спирохеты являются сапрофитами. Они встречаются на слизистой оболочке полости рта и половых органов здоровых людей.
Спирохеты могут образовывать цисты. Функция цисты - сохранение в неблагоприятных условиях (поэтому могут быть рецидивы). Не образуют спор, нет экзожгутиков.
Спирохеты не инертны - продуцируют ферменты и ферменты-токсины. В практических условиях биохимические методы не используются, но у неё есть сахаролитические, протеолитические ферменты. Имеют ферменты-токсины, например, гиалуронидаза.
Все заболевания, которые вызывают спирохеты, можно называть спирохетозы (трепонемозы, бореллозы, лептоспирозы).
Спирохетозы могут протекать остро и хронически. Могут поражаться животные, птицы. Для них характерен генерализованный характер, спирохетемия. Характерны рецидивы, либо течение в отдельных стадиях (сифилис). На некоторых стадиях спирохетозы дают кожные сыпи. Спирохеты устойчивы к пенициллину. Среди них есть антропонозы, зоонозы и антрозоонозы.
Спирохеты имеют извитую форму (есть и другие бактерии с извитой формой - вирионы, спириллы). За счёт чего вообще зависит та или иная форма, которую принимает бактерия? - От клеточной стенки, а точнее за счёт слоя пептидогликана. У спирохет пептидогликана очень мало, клеточная стенка легко поддаётся изменениям. Но кроме этого в цитоплазме бактериальной клетки находится фибриллярные нити (фибриллярный аппарат). Благодаря этому они и имеют извитую форму. Количество завитков, форма завитков у разных родов различна, поэтому микроскопический метод здесь является важным.
Спирохеты плохо воспринимают анилиновые красители (по Граму окрашиваются в красный цвет). Основная окраска - по Романовскому-Гимзы. При этом они могут раскрашиваться в разные цвета (красный, синий, фиолетовый). Лучше всего окрашиваются Borellia (синий), хуже Leptospira (красный) и ещё хуже - Treponema (слаборозовая).
Спирохеты - подвижные бактерии. Но движение здесь происходит не за счёт жгутиков, а за счёт сокращения осевой нити. А так как стенка ригидна, то передвижение таким образом идёт легко. Разные спирохеты могут передвигаться по-разному: ввинчиваться в среду, змеиться, плавно и т.д. (но принцип один). Так как есть движение, то исползуют нативный мазок. Метод микроскопии - темнопольный, может быть - фазово-контрастный.
Лучше всего они чувствуют себя в жидкой среде, в организме - на слизистых. По дыханию могут быть анаэробы, факультативные аэробы или аэробы. Они требовательны к питательной среде. Все спирохеты культивируют в жидкой или полужидкой среде, но со специальными свойствами. Только Leptospira может расти на обычных средах.Спирохеты всех видов выращивают на искусственных питательных средах. Основой питательной среды является сыворотка крови кролика или лошади, как цельная, так и разведенная изотоническим раствором хлорида натрия в различных соотношениях.
23. Впервые микоплазмы были обнаружены французскими ученьщи Нокаром и Ру в 1898 г. в фильтрате плевральной жидкости коров, больных плевропневмонией. Поэтому первоначально их назвали возбудителями плевропневмонии — РРО (pleuropneumonia organisms). В дальнейшем сходные с РРО организмы были найдены у человека и животных при различных патологических состояниях: заболеваниях ревматического характера, инфекциях дыхательных путей, воспалении мочеполовой системы. Все они были обозначены как PPLO — организмы, сходные с возбудителями плевропневмонии (pleuropneumonia like organisms). Микоплазмы имеют различную форму: сферическую, овальную, тонких нитей и звезд. По размерам большие из них приближаются к бактериям, мельчайшие (125— 150 нм) могут, как и вирусы, проходить через поры фарфоровых фильтров. Микоплазмы не имеют клеточной стенки и окружены тонкой трехслойной цитоплазматической мембраной, состоящей из липопротеидов. В отличие от вирусов они содержат как ДНК, так и РНК; их можно выращивать на искусственных питательных средах при добавлении лошадиной сыворотки. По классификации Берджи микоплазмы отнесены к группе 19. Микоплазмы широко распространены в природе. Их выделяют из почвы, сточных вод, от животных и человека. Известны микоплазмы, обитающие на слизистых оболочках рта и половых путей. Микоплазмы по своей морфологии и биологии сходны со стабильными L-формами бактерий. Поэтому предполагают, что микоплазмы возникли в результате генетических- мутаций из бактерий, лишившихся клеточной стенки.
L-формы — бактерии, частично или полностью лишённые клеточной стенки, но сохранившие способность к развитию.
Культивировать L-формы можно только на специальных средах препятствующих осмотическому разрушению клеток. L-формы лучше растут на плотной, чем в жидкой среде. На плотной среде они образуют колонии, врастающие в агар и имеющие характерную форму перевернутой шляпы. Колонии растут медленно, хотя иногда достигают значительных размеров.
24. Риккетсии представляют особую группу микроорганизмов, включающую как сапрофиты, так и патогенные виды. Название «риккетсии» они получили в честь американского ученого Риккетса, который впервые в 1910 г. обнаружил возбудителя сыпного тифа в крови больного и в зараженных вшах. Морфологически они сходны между собой, напоминают бактерии, полиморфны (могут иметь форму очень мелких палочек, кокков, нитей), грамотрицательны.
Как и вирусы, риккетсии являются внутриклеточными паразитами и размножаются только в клетках живого организма: в цитоплазме и ядре клеток.
Культивируют их в организме животного (морские свинки, белые мыши), насекомых-переносчиков (вши, клещи), в переживающей культуре ткани и курином эмбрионе ( в желточном мешке по Коксу). Устойчивость риккетсий особенно при нагревании, невелика: они гибнут при 80°С в течение минуты. Однако в высушенном состоянии, например в сухих фекалиях вши, сохраняются до 56 дней, а при 5°С — до 4 мес. Риккетсии являются возбудителями различных лихорадочных заболеваний, которые называются риккетсиозами. Они вызывают заболевай, клещи, блохи), диких и домашних животных и у человека. У вшей риккетсиоз заканчивается гибелью, у клещей и блох он протекает длительное время бессимптомно. Среди диких животных риккетсиозы поражают главным образом грызунов. Некоторые виды риккетсий вызывают заболевания у крупного и мелкого рогатого скота. У человека риккетсии вызывают различные заболевания. П. Ф. Здродовский разделяет риккетсиозы человека на пять групп:
I — группа сыпного тифа, включающая эпидемический и эндемический сыпной тиф;
II — группа клещевой пятнистой лихорадки, куда относятся:
1) пятнистая лихорадка Скалистых гор Америки;
2) марсельская, или средиземноморская, лихорадка;
3) североавстралийский клещевой сыпной тиф;
4) клещевой риккетсиоз, или клещевой сыпной тиф Средней Азии;
5) осповидный, или везикулярный, риккетсиоз;
III — группа краснотелково-клещевой лихорадки, или группа цуцугамуши, куда входит японская речная лихорадка цуцугамуши;
IV — группа пневмотропных риккетсиозов, или группа Ку- лихорадки; к ней относится одно заболевание — Ку-лихорадка;
V — группа пароксизмальных риккетсиозов, в которую входят:
1) волынская, или траншейная, лихорадка;
2) клещевой пароксизмальный риккетсиоз. Для большинства риккетсиозов характерна передача через кровососущих насекомых — вшей, блох, клещей, которые выделяют риккетсии с фекалиями (вши, блохи) или слюной (клещи). Реже риккетсиозы распространяются через выделения больных животных. Микробиологическая диагностика. Основана на выделении возбудителя при заражении чувствительных экспериментальных животных или куриных эмбрионов,. Эта методика требует наличия специальных лабораторий, известного навыка в работе и учете результатов. Поэтому основными методами диагностики риккетсиозов в практических лабораториях являются серологические исследования. Используют реакцию агглютинации со специфическими риккетсиозными антигенами, а такжеКРСк и РHA.
25. Правила работы в лаборатории
1. В помещение лаборатории нельзя входить без специальной одежды – халата.
2. Не разрешается выходить в халате за пределы лаборатории и надевать на халат верхнюю одежду.
3. В помещении лаборатории запрещается принимать пищу и хранить продукты питания.
4. Не выносить за пределы лаборатории, какие бы то ни было посуду и материалы, которые используются для проведения лабораторных работ (пробирки, краски и т. д.).
5. Не класть на стол личные вещи (сумки, папки и др.), держать их на специально отведенных местах.
6. Если микроорганизмы попадают на оборудование или пол (разобьется пробирка или чашка Петри, на которой они росли), об этом надо сразу же сообщить преподавателю или
лаборанту, а на данном месте провести обеззараживание, залив его дезинфицирующим раствором. После этого необходимо провести уборку.
7. Во время выполнения практических работ нельзя открывать форточки. Необходимо соблюдать тишину, избегать лишнего движения и хождения, открывания и закрывания дверей – всего того, что усиливает движение воздуха.
8. Перед началом работы дежурные проводят влажную уборку помещения, а столы протирают дезинфицирующим раствором.
9. Каждый студент перед началом работы должен проверить, все ли необходимое находится на его столе и исправен ли микроскоп.
10. Раздача необходимого для проведения лабораторной работы материала и посуды проводится лаборантом или дежурными.
11. На занятиях студенты должны иметь тетрадь и карандаши (простой и цветные – красный и синий). Рисунки при микроскопировании надо делать с препаратов, а не из книг или
пособий.
12. По окончании работы все используемые инструменты обеззараживают. Бактериальные петли и иглы прокаливают над пламенем спиртовки, а пипетки и стекла помещают в дезинфицирующий раствор.
13. Все используемые при работе микробные культуры сдают лаборанту, который проводит их обеззараживание или в автоклаве, или в дезинфицирующем растворе.
14. В конце занятий надо привести в порядок рабочий стол, протереть и убрать микроскоп, тщательно вымыть руки (при работе с заразным материалом их сначала дезинфицируют) и снять халат.
Уборка рабочего места
По окончании работы берут пинцетом кусок ваты, смачивают его в 5% растворе хлорамина или в 5% растворе формалина и протирают им поверхность стола на рабочем месте. Такого рода повседневная дезинфекция носит профилактический характер.
26. Вирусология как новая область инфекционной патологии возникла в конце XIX-го века, когда стало ясно, что многие распространенные заразные болезни человека, животных и растений вызываются иными возбудителями, чем бактерии и простейшие. Этими возбудителями оказались вирусы, впервые открытые Д.И. Ивановским (1892 г.) при изучении причины мозаичной болезни табака.
Строение и классификация вирусов
Вирусы относятся к царству Vira. Это
мельчайшие микробы («фильтрующиеся агенты»),
не имеющие клеточного строения, белоксинтезирующей системы,
содержащие один тип нуклеиновой кислоты (только ДНК или РНК).
Вирусы, являясь облигатными внутриклеточными паразитами, размножаются в цитоплазме или ядре клетки.
Они являются автономными генетическими структурами и отличаются особым, разобщенным (дизъюнктивным), способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы.
Сформированная вирусная частица называется вирионом.
Морфологию и структуру вирусов изучают с помощью электронной микроскопии, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Форма вирионов может быть различной (рис.):
палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), нитевидной (филовирусы), в виде сперматозоида (многие бактериофаги).
Различают ДНК- и РНК-содержащие вирусы. Исключением являются ретро-вирусы, имеющие диплоидный геном. Геном вирусов содержит от шести до нескольких сотен генов и представлен различными видами нуклеиновых кислот:
двунитевыми, однонитевыми, линейными, кольцевыми, фрагментированными.
Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную (геномную) функцию и функцию информационной РНК (иРНК).
Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Геном вирусов способен включаться в геном клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов, например, вирусов герпеса, могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды.
Различают:
просто устроенные вирусы (например, вирусы полиомиелита, гепатита А) и
сложно устроенные вирусы (например, вирусы кори, гриппа, герпеса, коронавирусы).
У просто устроенных вирусов (рис.) нуклеиновая кислота связана с белковой оболочкой, называемой капсидом (от лат. capsa—футляр). У сложноустроенных вирусов (рис.) капсид окружен липопротеиновой оболочкой — суперкапсидом, или пеплосом.
В основу классификации вирусов положены следующие категории:
тип нуклеиновой кислоты (ДНК или РНК), ее структура, количество нитей (одна или две), особенности воспроизводства вирусного генома (табл. 2.3),
размер и морфология вирионов, количество капсомеров и тип симметрии нуклеокапсида, наличие оболочки (суперкапсида).
чувствительность к эфиру и дезоксихолату,
место размножения в клетке,
антигенные свойства и др.
разница между вирусами и бактериями заключается в следующем:
Вирусы являются доклеточными живыми организмами, в то время как бактерии – одноклеточные.
Вирусы не могут размножаться вне живых клеток. Бактерии размножаются самостоятельно делением клетки.
Вирусы обладают только одним видом нуклеиновой кислоты, а в состав бактерии входит ДНК, РНК, рибосомы и клеточная мембрана.
27. Непременным компонентом вирусной частицы является одна из двух нуклеиновых кислот, белок и зольные элементы. Эти три компонента являются общими для вирусов, тогда как остальные двалипоиды и углеводы - входят в состав далеко не всех вирусов.
Вирусы, состоящие только из белка нуклеиновой кислоты и зольных элементов, чаще всего принадлежат к группе простых вирусов, лишенных дифференциации, собственных ферментов или каких-либо специализированных структур – вирусы растений, некоторые вирусы животных и насекомых. В то же время практически все бактериофаги, которые по химическому составу, принадлежат к группе минимальных вирусов, на самом деле являются очень сложными и
высокодифференцированными структурами. Вирусы, в состав которых наряду с белком и нуклеиновой кислотой входят также липоиды и углеводы, как правило, принадлежат к группе сложно устроенных вирусов. Большая часть вирусов этой группы паразитирует на животных.
Белки вирусов
Белок всех исследованных до настоящего времени вирусов построен из обычных аминокислот, принадлежащих к естественному L-ряду. Соотношение аминокислот в вирусных белках достаточно близко к таковому в белках животных, бактерий и растений. Вирусные белки не содержат обычно большого количества основных аминокислот (аргинина, муцина). Не учитывая нейтральных аминокислот, можно сказать, что в вирусном белке преобладают кислые дикарбоновые кислоты. Это справедливо для вирусов с низким и высоким содержанием нуклеиновых кислот.
Вирусная ДНК
Молекулы вирусных ДНК могут быть линейными или кольцевыми, двух цепочечными или одно цепочечными по всей своей длине или же одно цепочечными только на концах. Кроме того, выяснилось, что большинство нуклеотидных последовательностей в вирусном геноме встречается лишь по одному разу, однако на концах могут находиться повторяющиеся, или избыточные участки. Помимо различий в форме молекулы и в структуре концевых участков вирусных ДНК
существуют также различия в величине генома.
Вирусная РНК
Исследования вирусной РНК составили один из самых значительных вкладов вирусологии в молекулярную биологию. Тот факт, что у вирусов растений реплицируемая генетическая система состоит только из РНК, ясно показал, что и РНК способна сохранять генетическую информацию. Была установлена инфекционность РНК вируса табачной мозаики, и выяснилось, что для инфекции
необходима вся ее молекула. Размеры вирионов РНК - вирусов сильно варьируют - от 7.106
до 2.108 дальтон, однако размеры РНК и, следовательно, объем содержащейся в ней информации различаются в значительно меньшей степени. Основные методы культивирования вирусов:
1) биологический – заражение лабораторных животных. При заражении вирусом животное заболевает. Если болезнь не развивается, то патологические изменения можно обнаружить при вскрытии. У животных наблюдаются иммунологические сдвиги. Однако далеко не все вирусы можно культивировать в организме животных;
2) культивирование вирусов в развивающихся куриных эмбрионах. Куриные эмбрионы выращивают в инкубаторе 7—10 дней, а затем используют для культивирования. В этой модели все типы зачатков тканей подвержены заражению. Но не все вирусы могут размножаться и развиваться в куриных эмбрионах.
В результате заражения могут происходить и появляться:
1) гибель эмбриона;
2) дефекты развития: на поверхности оболочек появляются образования – бляшки, представляющие собой скопления погибших клеток, содержащих вирионы;
3) накопление вирусов в аллантоисной жидкости (обнаруживают путем титрования);
4) размножение в культуре ткани (это основной метод культивирования вирусов).
Различают следующие типы культур тканей:
1) перевиваемые – культуры опухолевых клеток; обладают большой митотической активностью;
2) первично трипсинизированные – подвергшиеся первичной обработке трипсином; эта обработка нарушает межклеточные связи, в результате чего выделяются отдельные клетки. Источником являются любые органы и ткани, чаще всего – эмбриональные (обладают высокой митотической активностью).
Для поддержания клеток культуры ткани используют специальные среды. Это жидкие питательные среды сложного состава, содержащие аминокислоты, углеводы, факторы роста, источники белка, антибиотики и индикаторы для оценки развития клеток культуры ткани.
28. Ви́рус (лат. virus — яд) — субклеточный инфекционный агент, который может воспроизводиться только внутри живых клеток организма. Они поражают все типы организмов, от растений и животных до бактерий и архей[3] (вирусы бактерий обычно называют бактериофагами).
Отличительные признаки:
1) содержат лишь один тип нуклеиновой кислоты (РНК или ДНК);
2) не имеют собственных белоксинтезирующих и энергетических систем;
3) не имеют клеточной организации;
4) обладают дизъюнктивным (разобщенным) способом репродукции (синтез белков и нуклеиновых кислот происходит в разных местах и в разное время);
5) облигатный паразитизм вирусов реализуется на генетическом уровне;
6) вирусы проходят через бактериальные фильтры.
Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитоли-тическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
проходит в 6 стадий:
1) адсорбция вирионов на клетке;
2) проникновение вируса в клетку;
3) «раздевание» и высвобождение вирусного генома (депротеинизация вируса);
4) синтез вирусных компонентов;
5) формирование вирионов;
6) выход вирионов из клетки.
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас- плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты: РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус- нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов. Этот тип взаимодействия не завершается образованием вирусного потомства и может возникать при следующих обстоятельствах:
1 ) заражение
чувствительных клеток дефектными
вирусами или дефектными вирионами;
) заражение
чувствительных клеток дефектными
вирусами или дефектными вирионами;
заражение стандартным вирусом генетически резистентных к нему клеток;
заражение стандартным вирусом чувствительных клеток в непермиссивных (неразрешающих) условиях.
Интегративный тип, или вирогения —характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Интегративный тип взаимодействия характерен:
для умеренных ДНК-содержащих бактериофагов,
онкогенных вирусов
и некоторых инфекционных вирусов как ДНК-содержащих (например, вируса гепатита В), так и РНК-содержащих (например, вируса иммунодефицита человека).
Для интеграции с геномом клетки необходимо наличие кольцевой формы двунитевой ДНК-вируса. Геном ДНК-содержащих вирусов в кольцевой форме прикрепляется к клеточной ДНК в месте гомологии нуклеотидных последовательностей и встраивается в определенный участок хромосомы при участии ряда ферментов (рестриктаз, эндонуклеаз, лигаз).
У РНК-содержащих вирусов процесс интеграции более сложный. Он начинается с механизма обратной транскрипции, который заключается в синтезе комплементарной нити ДНК на матрице вирусной РНК с помощью вирусоспецифического фермента обратной транскриптазы (ревертазы). После образования двунитевой ДНК и замыкания ее в кольцо происходит интеграция ДНК-транскрипта в хромосому клетки.
29. Методы микробиологической диагностики вирусных инфекций
В вирусологии методы лабораторной диагностики вирусных инфекций имеют свою специфику, учитывая особенности биологии вирусов. Используются вирусоскопический, вирусологический и серологический методы лабораторной диагностики.
Вирусоскопический метод заключается в обнаружении вируса в исследуемом материале под микроскопом. Чаще используют электронный микроскоп, реже - люминесцентный. Световая микроскопия из-за ничтожно малых размеров вирусов практически не применяется. Лишь для обнаружения крупных вирусов, используя методы сверхокраски, можно применить световой микроскоп. Кроме того, с помощью светового микроскопа можно выявить внутриклеточные включения, которые образуются в пораженных клетках при некоторых инфекциях.
Вирусологический метод заключается в заражении исследуемым материалом чувствительной биологической модели (лабораторные животные, куриные эмбрионы или культуры клеток), индикации вируса и его последующей идентификации. При заражении лабораторных животных индикация вирусов производится, как правило, по клинической картине болезни, патолого-анатомическим изменениям ориентировочно и окончательно, например, с помощью реакции гемагглютинации. Эта же реакция позволяет выявить вирусы в курином эмбрионе, видимых изменений при вскрытии которого, как правило, не наблюдается. В культуре клеток наличие вируса определяют по цитопатическому действию (в том числе образованию внутриклеточных включений), гемадсорбции, феномену бляшкообразования, реакции гемагглютинации, отсутствию изменения окраски индикатора. Идентификация вируса осуществляется с помощью серологических реакций (РПГА, РТГА, РН, РСК, ИФА и др.). Вирусологический метод позволяет точно определить природу возбудителя, но он требует достаточного времени (5-7 дней и более), значительных материальных затрат и небезопасен.
|
Особенностью серологического метода в вирусологии является исследование парных сывороток. Первую сыворотку берут у больного в острый период в начале болезни, хранят при температуре 4-8 ?С, а вторую сыворотку берут через 10-14 дней. Сыворотки
исследуют одномоментно. О болезни свидетельствует сероконверсия, т.е. нарастание титра антител во второй сыворотке по отношению к первой. Диагностической является сероконверсия в 4 раза и выше. Так как многие вирусные болезни протекают остро, этот вариант серологического метода обычно применяют для ретроспективной диагностики.
Ведущим методом лабораторной диагностики вирусных инфекций является вирусологический.
Ускоренная и экспресс-диагностика вирусных болезней производится так же, как при бактериальных инфекциях.
30 Методы культивирования вирусов.
Для культивирования вирусов используют культуры клеток, куриные
эмбрионы и чувствительных лабораторных животных. Эти же методы
используют и для культивирования риккетсий и хламидий - облигатных
внутриклеточных бактерий, которые не растут на искусственных питательных
средах.
Культуры клеток. Культуры клеток готовят из тканей животных или
человека. Культуры подразделяют на первичные (неперевиваемые),
полуперевиваемые и перевиваемые.
Приготовление первичной культуры клеток складывается из нескольких
последовательных этапов: измельчения ткани, разъединения клеток путем
трипсинизации, отмывания полученной однородной суспензии изолированных
клеток от трипсина с последующим суспендированием клеток в питательной
среде, обеспечивающей их рост, например в среде 199 с добавлением телячьей
сыворотки крови.
Перевиваемые культуры в отличие от первичных адаптированы к условиям,
обеспечивающим им постоянное существование in vitro, и сохраняются на
протяжении нескольких десятков пассажей.
Перевиваемые однослойные культуры клеток приготовляют из
злокачественных и нормальных линий клеток, обладающих способностью
длительно размножаться in vitro в определенных условиях. К ним относятся
злокачественные клетки HeLa, первоначально выделенные из карциномы
шейки матки, Нер-3 (из лимфоидной карциномы), а также нормальные клетки
амниона человека, почек обезьяны и др.
К полуперевиваемым культурам относятся диплоидные клетки человека. Они
представляют собой клеточную систему, сохраняющую в процессе 50
пассажей (до года) диплоидный набор хромосом, типичный для соматических
клеток используемой ткани. Диплоидные клетки человека не претерпевают
злокачественного перерождения и этим выгодно отличаются от опухолевых.
О размножении (репродукции) вирусов в культуре клеток судят по
цитопатическому действию (ЦПД), которое может быть обнаружено
микроскопически и характеризуется морфологическими изменениями клеток.
Характер ЦПД вирусов используют как для их обнаружения (индикации), так и
для ориентировочной идентификации, т. е. определения их видовой
принадлежности.
Один из методов индикации вирусов основан на способности поверхности
клеток, в которых они репродуцируются, адсорбировать эритроциты -
реакция гемадсорбции. Для ее постановки в культуру клеток, зараженных
вирусами, добавляют взвесь эритроцитов и после некоторого времени контакта
клетки промывают изотоническим раствором хлорида натрия. На поверхности
пораженных вирусами клеток остаются прилипшие эритроциты.
Другой метод - реакция гемагглютинации (РГ). Применяется для
обнаружения вирусов в культуральной жидкости культуры клеток либо
хорионаллантоисной или амниотической жидкости куриного эмбриона.
Количество вирусных частиц определяют методом титрования по ЦПД в
культуре клеток. Для этого клетки культуры заражают десятикратным
разведением вируса. После 6-7-дневной инкубации их просматривают на
наличие ЦПД. За титр вируса принимают наибольшее разведение, которое
вызывает ЦПД в 50 % зараженных культур. Титр вируса выражают
количеством цитопатических доз.
Более точным количественным методом учета отдельных вирусных частиц
является метод бляшек.
Некоторые вирусы можно обнаружить и идентифицировать по включениям,
которые они образуют в ядре или цитоплазме зараженных клеток.
Куриные эмбрионы. Куриные эмбрионы по сравнению с культурами
клеток значительно реже бывают контаминированы вирусами и микоплазмами,
а также обладают сравнительно высокой жизнеспособностью и устойчивостью
к различным воздействиям.
Для получения чистых культур риккетсий, хламидий. и ряда вирусов в
диагностических целях, а также для приготовления разнообразных препаратов
(вакцины, диагностикумы) используют 8-12-дневные куриные эмбрионы. О
размножении упомянутых микроорганизмов судят по морфологическим из-
менениям, выявляемым после вскрытия эмбриона на его оболочках.
О репродукции некоторых вирусов, например гриппа, оспы, можно судить по
реакции гемагглютинации (РГА) с куриными или другими эритроцитами.
К недостаткам данного метода относятся невозможность обнаружения
исследуемого микроорганизма без предварительного вскрытия эмбриона, а
также наличие в нем большого количества белков и других соединений,
затрудняющих последующую очистку риккетсий или вирусов при
изготовлении различных препаратов.
Лабораторные животные. Видовая чувствительность животных к
определенному вирусу и их возраст определяют репродуктивную способность
вирусов. Во многих случаях только новорожденные животные чувствительны
к тому или иному вирусу (например, мыши-сосунки - к вирусам Коксаки).
Преимущество данного метода перед другими состоит в возможности
выделения тех вирусов, которые плохо репродуцируются в культуре или
эмбрионе. К его недостаткам относятся контаминация организма подопытных
животных посторонними вирусами и микоплазмами, а также необходимость
последующего заражения культуры клеток для получения чистой линии данно-
го вируса, что удлиняет сроки исследования.
|
31 вопрос Бактериофаги (от бактерии и греч. phagos — пожиратель; буквально — пожиратели бактерий), фаги, бактериальные вирусы, вызывающие разрушение (лизис) бактерий и других микроорганизмов. Б. размножаются в клетках, лизируют их и переходят в др., как правило, молодые, растущие клетки. Впервые перевиваемый лизис бактерий (сибиреязвенной палочки) наблюдал в 1898 русский микробиолог Н. Ф. Гамалея. В 1915 английский учёный Ф. Туорт описал это же явление у гнойного стафилококка, а в 1917 французский учёный Ф. Д'Эрелль назвал литический агент, проходящий через бактериальные фильтры, "Б".Строение и химический состав. Частицы многих Б. состоят из головки округлой, гексагональной или палочковидной формы диаметром 45—140 нм и отростка толщиной 10—40 и длиной 100—200 нм (рис.). Другие Б. не имеют отростка; одни из них округлы, другие — нитевидны, размером 8х800 нм. Содержимое головки состоит преимущественно из дезоксирибону клейновой кислоты (ДНК) (длина её нити во много раз превышает размер головки и достигает 60—70 мкм, эта нить плотно скручена в головке) или рибонуклеиновой кислоты (РНК) и небольшого количества (около 3%) белка и некоторых других веществ. Отросток имеет вид полой трубки, окруженной чехлом, содержащим сократительные белки, подобные мышечным. У ряда Б. чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих Б. имеется базальная пластинка с несколькими шиловидными или другие формы выступами. От пластинки отходят тонкие длинные нити, которые способствуют прикреплению фага к бактерии (см. схему). Оболочки головки и отростка состоят из белков. Общее количество белка в частице фага 50—60% , нуклеиновых кислот — 40—50% . Каждый Б. обладает специфическими антигенными свойствами, отличными от антигенов бактерии-хозяина и других фагов. Имеются антигены, общие для ряда фагов (особенно содержащих РНК).Распространение. Б. найдены для большинства бактерий, в том числе патогенных и сапрофитных, а также .для актиномицетов (актинофаги) и сине-зелёных водорослей. Встречаются Б. в кишечнике человека и животных, в растениях, почве, водоёмах, сточных водах, навозе и т. д. Б. почвенных микроорганизмов влияют на течение микробиологических процессов в почве (денитрификацию, аммонификацию, азотфиксацию). Размножение. Б. прикрепляется своим отростком к бактериальной клетке и, выделяя фермент, растворяет клеточную стенку; затем содержимое его головки через канадец отростка переходит внутрь клетки, где под влиянием нуклеиновой кислоты фага останавливается синтез бактериальных белков, ДНК и РНК и начинается синтез нуклеиновой кислоты, а затем и белков фага. Часть этих белков — ферменты, другая часть образует оболочку зрелой частицы Б. Более мелкие, сферические фаги попадают в бактерии без участия отростка. Если клетка бактерии заражена одновременно частицами Б., различающимися между собой по ряду свойств, то среди потомства, кроме частиц, подобных родителям, будут и такие, у которых эти свойства встречаются в новой комбинации, т. к. при размножении Б. наблюдается рекомбинация — обмен кусками нитей нуклеиновой кислоты, являющейся носителем наследственной информации. Частицы крупных фагов выходят из бактерии, разрушая её, а некоторых мелких и нитевидных — из живых бактерий (см. Вирусы). Одни Б. весьма специфичны и способны лизировать клетки только одного какого-либо вида микроорганизмов (монофаги), другие — клетки разных видов (полифаги). Б. делят на вирулентные, вызывающие лизис клетки с образованием новых частиц, и умеренные (симбиотические), которые адсорбируются клеткой и проникают в неё, но лизиса не вызывают, а остаются в клетке в латентной (скрытой) неинфекционной форме (профаг). Культуры, содержащие латентный фаг, называются лизогенными. Лизогения передаётся потомству бактерии. Лизогенная культура может содержать 2—3 и более фагов; она, как правило, устойчива против находящихся в ней фагов (лишь небольшая часть клеток лизируется и освобождает зрелые фаги). Воздействуя на лизогенную культуру ультрафиолетовыми или рентгеновскими лучами, перекисью водорода и некоторыми другими веществами, можно значительно увеличить количество клеток, освобождающих фаг (т. н. индукция Б.). Лизогения широко распространена среди всех видов бактерий и актиномицетов. В ряде случаев многие свойства лизогенной культуры (токсичность, подвижность бактерий и др.) зависят от наличия в ней определённых профагов. Описано много мутаций Б., сопровождающихся изменением их литической активности, строения частиц и "колоний", устойчивости против неблагоприятных воздействий и другие свойств. Б. играют большую роль в изменчивости и эволюции микробов, причём механизмы воздействия их на клетку разные (см. Трансдукция). Б. могут резко изменять азотфиксирующую способность азотобактера, токсичность и антигенные свойства патогенных бактерий и др. Практическое значение Б. Некоторые фаги (одни или в сочетании с антибиотиками) применяли для профилактики (фагопрофилактики) и лечения (фаготерапии) ряда бактериальных инфекционных болезней человека (дизентерия, брюшной тиф, холера, чума, стафилококковые и анаэробная инфекции и др.) и животных. Однако антибиотики и другие химиотерапевтические средства оказались эффективнее фагов, в связи с чем применение их с лечебной целью сузилось. Б. успешно применяются при определении вида бактерий, актиномицетов. Б. могут вредить производству антибиотиков, аминокислот, молочных продуктов, бактериальных удобрений и в других отраслях микробиологического синтеза. Велико значение Б. для теоретических работ по генетике и молекулярной биологии. |
32,33Бактериофаги. Взаимодействие фага с бактериальной клеткой.
Умеренные и вирулентные бактериофаги. Лизогения.
Бактериофаги - вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, репродуцироваться в них и вызывать их растворение (лизис).
Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги. Вирулентные фаги, проникнув в бактериальную клетку, автономно репродуцируются в ней и вызывают лизис бактерий. Процесс взаимодействия вирулентного фага с бактерией протекает в виде нескольких стадий и весьма схож с процессом взаимодействия вирусов человека и животных с клеткой хозяина. Однако для фагов, имеющих хвостовой отросток с сокращающимся
чехлом, он имеет особенности. Эти фаги адсорбируются на поверхности бактериальной клетки с помощью фибрилл хвостового отростка. В результате активации фагового фермента АТФазы происходит сокращение чехла хвостового отростка и внедрение стержня в клетку. В процессе "прокалывания" клеточной стенки бактерии принимает участие фермент лизоцим, находящийся на конце хвостового отростка. Вслед за этим ДНК фага, содержащаяся в головке, проходит через полость хвостового стержня и ак-
тивно впрыскивается в цитоплазму клетки. Остальные структурные элементы фага (капсид и отросток) остаются вне клетки. После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фаговоголизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30-40 мин. Процесс бактериофагии проходит несколько циклов, пока не будут лизированы все чувствительные к данному фагу бактерии.
Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий.
Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков.
Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии.
Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов.
34 Применение фагов в биотехнологии, микробиологии и медицине.
Практическое применение фагов. Бактериофаги используют в лабораторной
диагностике инфекций при внутривидовой идентификации бактерий, т. е.
определении фаговара (фаготипа). Для этого применяют метод
фаготипирования, основанный на строгой специфичности действия фагов: на
чашку с плотной питательной средой, засеянной "газоном" чистой культурой
возбудителя, наносят капли различных диагностических типоспецифических
фагов. Фаговар бактерии определяется тем типом фага, который вызвал ее
лизис (образование стерильного пятна, "бляшки", или "негативной колонии",
фага). Методику фаготипирования используют для выявления источника и
путей распространения инфекции (эпидемиологическое маркирование).
Выделение бактерий одного фаговара от разных больных указывает на общий
источник их заражения.
По содержанию бактериофагов в объектах окружающей среды (например, в
воде) можно судить о присутствии в них соответствующих патогенных
бактерий. Подобные исследования проводят при эпидемиологическом анализе
вспышек инфекционных болезней.
Фаги применяют также для лечения и профилактики ряда бактериальных
инфекций. Производят брюшнотифозный, сальмонеллезный, дизентерийный,
синегнойный, стафилококковый, стрептококковый фаги и комбинированные
препараты (колипротейный, пиобактериофаги и др). Бактериофаги назначают
по показаниям перорально, парентерально или местно в виде жидких, таблети-
рованных форм, свечей или аэрозолей.
Бактериофаги широко применяют в генной инженерии и биотехнологии в
качестве векторов для получения рекомбинантных ДНК.