

В
ходе различных исследований было
выяснено, что большинство генов эукариот
оказалось разорванными (split) на несколько
кодирующих участков (экзонов), разделенных
находящимися между ними некодирующими
последовательностями - интронами.
Оказалось, что внутри генов эукариот
всегда бывают участки, которые не
кодируют ни полипептидов, ни стабильных
РНК. Эти участки назвали интронами —
от английских слов intervening zone — зона,
„перемежающая“ смысловую последовательность
гена. А кодирующие участки были названы
экзонами (от expressing zone — экспрессируемая
зона гена). Во многих случаях каждому
экзону соответствует домен — часть
белковой молекулы, обладающая относительной
завершённостью и независимостью.
 Интрон
— участок ДНК, который является
частью гена,
но не содержит информации о
последовательности аминокислот белка.
Иными словами, это транскрибируемый участок ДНК,
который удаляется из состава транскипта
при сплайсинге;
в результате последовательности,
находящиеся по обе стороны от интрона
(экзоны) объединяются.
Последовательностьнуклеотидов,
соответствующая интрону, удаляется из
транскрибированной с него мРНК в
процессе сплайсинга до
того, как произойдет считывание белка
(трансляция).
Интрон
— участок ДНК, который является
частью гена,
но не содержит информации о
последовательности аминокислот белка.
Иными словами, это транскрибируемый участок ДНК,
который удаляется из состава транскипта
при сплайсинге;
в результате последовательности,
находящиеся по обе стороны от интрона
(экзоны) объединяются.
Последовательностьнуклеотидов,
соответствующая интрону, удаляется из
транскрибированной с него мРНК в
процессе сплайсинга до
того, как произойдет считывание белка
(трансляция).
 Интроны
характерны для всех типов эукариотической РНК,
но также найдены в рибосомальной
РНК (рРНК) и транспортной
РНК (тРНК) прокариот.
Число и длина интронов очень различны
в разных видах и среди разных генов
одного организма. Гены млекопитающих,
в том числе человека, и цветковых растений
часто содержат многочисленные интроны,
которые могут быть длиннее экзонов.
Кодирующие последовательности, которые
в конечном итоге и дают мРНК, показаны
черным, а интроны - белым:
Интроны
характерны для всех типов эукариотической РНК,
но также найдены в рибосомальной
РНК (рРНК) и транспортной
РНК (тРНК) прокариот.
Число и длина интронов очень различны
в разных видах и среди разных генов
одного организма. Гены млекопитающих,
в том числе человека, и цветковых растений
часто содержат многочисленные интроны,
которые могут быть длиннее экзонов.
Кодирующие последовательности, которые
в конечном итоге и дают мРНК, показаны
черным, а интроны - белым:
 Число
интронов достигает иногда поразительно
высоких значений, если учесть
точность процессинга,
необходимого для продуцирования
функциональной мРНК: 33 интрона для
гена вителлогенина
A(Wahli
et al.) и
даже 50 интронов для коллагенового гена (de
Crombrugghe et al.).
Число
интронов достигает иногда поразительно
высоких значений, если учесть
точность процессинга,
необходимого для продуцирования
функциональной мРНК: 33 интрона для
гена вителлогенина
A(Wahli
et al.) и
даже 50 интронов для коллагенового гена (de
Crombrugghe et al.).
2
Классификация интронов
 Существует
четыре группы интронов:
Ядерные
интроны
Интроны
группы I
Интроны
группы II
Интроны
группы III
Иногда
интроны группы III также относят к группе
II, потому что они похожи по структуре и
функции.
Ядерные, или спалайсосомные
интроны подвергаются сплайсингу при
помощи сплайсосомы и малых
ядерных РНК (мяРНК,
snRNA). В последовательности РНК, содержащей
ядерные интроны, есть специальные
сигнальные последовательности, которые
узнаются сплайсосомой.
Интроны
I, II и III группы способны к автосплайсингу
и встречаются реже, чем сплайсосомные
интроны.
Существует
четыре группы интронов:
Ядерные
интроны
Интроны
группы I
Интроны
группы II
Интроны
группы III
Иногда
интроны группы III также относят к группе
II, потому что они похожи по структуре и
функции.
Ядерные, или спалайсосомные
интроны подвергаются сплайсингу при
помощи сплайсосомы и малых
ядерных РНК (мяРНК,
snRNA). В последовательности РНК, содержащей
ядерные интроны, есть специальные
сигнальные последовательности, которые
узнаются сплайсосомой.
Интроны
I, II и III группы способны к автосплайсингу
и встречаются реже, чем сплайсосомные
интроны.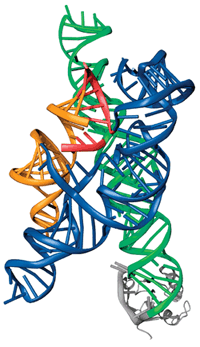 На
рисунке представлена схема кристаллической
структуры автосплайсинга интрона группы
I (экзон обозначен красным). Интроны II и
III группы похожи друг на друга и обладают
консервативной вторичной структурой.
Они обладают свойствами, похожими на
свойства сплайсосомы и, вероятно,
являются её эволюционными предшественниками.
Интроны I группы, которые встречаюся
у бактерий и простейших —
единственный класс интронов, который
требует присутствие несвязанного
гуанилового нуклеотида. Их вторичная
структура отличается от вторичной
структуры интронов II и III группы.
На
рисунке представлена схема кристаллической
структуры автосплайсинга интрона группы
I (экзон обозначен красным). Интроны II и
III группы похожи друг на друга и обладают
консервативной вторичной структурой.
Они обладают свойствами, похожими на
свойства сплайсосомы и, вероятно,
являются её эволюционными предшественниками.
Интроны I группы, которые встречаюся
у бактерий и простейших —
единственный класс интронов, который
требует присутствие несвязанного
гуанилового нуклеотида. Их вторичная
структура отличается от вторичной
структуры интронов II и III группы.
3
Генетический мусор и не только. Эволюционная и другие функции интронов
Существуют
две альтернативные теории, объясняющие
происхождение и эволюцию сплайсосомных
интронов: так называемые теории ранних
интронов (РИ), которую высказал Джилберт
(Gilbert),
и поздних интронов (ПИ). Теория РИ
утверждает, что многочисленные интроны
присутствовали в общих предках эу- и
прокариот и, соответственно, интроны
являются очень старыми структурами.
Согласно этой модели, интроны были
потеряны из генома прокариот. Также она
предполагает, что ранние интроны
способствовали рекомбинации экзонов,
представляющих домены белков. ПИ
утверждает, что интроны появились в
генах относительно недавно, и были
инсертированы (вставлены) в геном после
разделения организмов на про- и эукариоты.
Эта модель основывается на наблюдении,
что сплайсосомные интроны есть только
у эукариот.
Предположение об
эволюционной функции разорванных генов
высказал Джилберт (Gilbert).
Интронами часто разделяются кодирующие
последовательности, с которых транслируются
функциональные домены внутри белков.
Так, например, глобиновые гены
разделены двумя интронами на три
кодирующих участка. Центральный участок
кодирует домен,
связывающий гем.
Если, как полагает Джилберт, интроны
создают возможность для «перетасовки
ДНК путем незаконных рекомбинаций», то
в таком случае кодирующие последовательности
для отдельных доменов могут вступать
в новые комбинации друг с другом. Так,
участок глобина, связывающий гем,
возможно, первоначально составлял часть
другого разорванного гена. Для эволюции
будущего глобинового гена, возможно,
не понадобилось дупликации гена и
дивергенции: простой перетасовки
существующих доменов могло оказаться
достаточным для создания нового белка
из уже существовавших частей.
 Лизоцимы фага Т4
и куриного яйца содержат структуры, к
которым по мнениюАртымюка
и др. применимо
подобного рода эволюционное объяснение.
Лизоцимы куриного яйца содержат два
таких же домена как в лизоциме фага
Т4. Один
из них - это домен, содержащий каталитический
центр, а другой - соседний - домен,
по-видимому, участвует в определении
субстратной специфичности.
N-концевой домен лизоцима куриного яйца,
который, как
установили Янг и др. ,
содержит сигнальный пептид прелизоцима
и начальные аминокислоты молекул зрелого
белка, не имеет эквивалента в лизоциме
фага Т4. Точно так же четвертые, С-концевые,
домены этих двух белков очень сильно
различаются; по
мнению Мэтьюза и др. ,
в фаговом ферменте функция этого домена
состоит в том, чтобы обеспечивать
прикрепление фаговой частицы к
стенке Escherichia
coli,
тогда как ферменту куриного яйца эта
функция вряд ли необходима. Подобный
характер строения гомологичных белков,
когда к сходным доменам примыкают
несходные, явно совместим с предположением
о перетасовке соответствующих кодирующих
последовательностей путем рекомбинации
в пределах интронов.
Джилберт
высказал мнение, что если функция
интронов сводится к обеспечению
эволюционной пластичности, то они могут
утрачиваться в результате нейтрального
дрейфа. Он считает, что это происходит
с исключительно низкой скоростью. Если
согласиться с тем, что длительная
эволюционная пластичность служит
достаточным механизмом для сохранения
разорванных генов, то старая теория о
старении расы, к которой так часто
прибегали в конце XIX в., чтобы объяснить
вымирание различных групп (от аммонитов
до динозавров), может быть возрождена
в новой форме. Постаревшими будут
считаться те группы, которые в результате
дрейфа потеряли так много интронов, что
утратили способность противостоять
давлению отбора путем возникновения
эволюционных новшеств. Однако здесь мы
сталкиваемся с той же самой логической
ошибкой, которая была присуща старой
теории преформизма:
все интроны должны были присутствовать
изначально, а затем запас их должен
медленно истощаться. Представляется
более вероятным, что новые интроны могут
возникать и что их сохранение связано
с какой-то непосредственной функцией,
выполняемой ими в клетке. Это не означает,
однако, что отрицается возможность
дополнительной эволюционной роли
интронов в перетасовке ДНК.
Важную
роль интронов в регуляции генной
экспрессии продемонстрировали Лазовска,
Жак и Слонимски (Lazowska, Jacq, Slonimski) в
своем исследовании гена
box,
который локализован в митохондриальном
геноме дрожжей и кодирует цитохром b.
Физическая карта гена box и кластеров
известных для него мутационных сайтов
представлены на рисунке
Лизоцимы фага Т4
и куриного яйца содержат структуры, к
которым по мнениюАртымюка
и др. применимо
подобного рода эволюционное объяснение.
Лизоцимы куриного яйца содержат два
таких же домена как в лизоциме фага
Т4. Один
из них - это домен, содержащий каталитический
центр, а другой - соседний - домен,
по-видимому, участвует в определении
субстратной специфичности.
N-концевой домен лизоцима куриного яйца,
который, как
установили Янг и др. ,
содержит сигнальный пептид прелизоцима
и начальные аминокислоты молекул зрелого
белка, не имеет эквивалента в лизоциме
фага Т4. Точно так же четвертые, С-концевые,
домены этих двух белков очень сильно
различаются; по
мнению Мэтьюза и др. ,
в фаговом ферменте функция этого домена
состоит в том, чтобы обеспечивать
прикрепление фаговой частицы к
стенке Escherichia
coli,
тогда как ферменту куриного яйца эта
функция вряд ли необходима. Подобный
характер строения гомологичных белков,
когда к сходным доменам примыкают
несходные, явно совместим с предположением
о перетасовке соответствующих кодирующих
последовательностей путем рекомбинации
в пределах интронов.
Джилберт
высказал мнение, что если функция
интронов сводится к обеспечению
эволюционной пластичности, то они могут
утрачиваться в результате нейтрального
дрейфа. Он считает, что это происходит
с исключительно низкой скоростью. Если
согласиться с тем, что длительная
эволюционная пластичность служит
достаточным механизмом для сохранения
разорванных генов, то старая теория о
старении расы, к которой так часто
прибегали в конце XIX в., чтобы объяснить
вымирание различных групп (от аммонитов
до динозавров), может быть возрождена
в новой форме. Постаревшими будут
считаться те группы, которые в результате
дрейфа потеряли так много интронов, что
утратили способность противостоять
давлению отбора путем возникновения
эволюционных новшеств. Однако здесь мы
сталкиваемся с той же самой логической
ошибкой, которая была присуща старой
теории преформизма:
все интроны должны были присутствовать
изначально, а затем запас их должен
медленно истощаться. Представляется
более вероятным, что новые интроны могут
возникать и что их сохранение связано
с какой-то непосредственной функцией,
выполняемой ими в клетке. Это не означает,
однако, что отрицается возможность
дополнительной эволюционной роли
интронов в перетасовке ДНК.
Важную
роль интронов в регуляции генной
экспрессии продемонстрировали Лазовска,
Жак и Слонимски (Lazowska, Jacq, Slonimski) в
своем исследовании гена
box,
который локализован в митохондриальном
геноме дрожжей и кодирует цитохром b.
Физическая карта гена box и кластеров
известных для него мутационных сайтов
представлены на рисунке
 В
этом гене имеется шесть кодирующих
последовательностей и пять интронов;
у него обнаружены три различных класса
мутаций. Мутации в кодирующих
последовательностях, как и следовало
ожидать, влияют на строение белка, и все
они входят в одну группу комплементации.
Мутации двух других классов необычны.
Три кластера локализованы в интронах.
Они в свою очередь образуют три различные
группы комплементации и блокируют
процессинг транскрипта гена цитохрома
b. Они оказывают также воздействие на
экспрессию гена oxi-3 - еще одного
разорванного митохондриального гена,
который кодирует субъединицу
1 цитохромоксидазы.
Мутации третьего класса локализованы
на границах между интронами и кодирующими
последовательностями.
Для того
чтобы могла образоваться функциональная
мРНК для цитохрома b, интронные
последовательности должны быть вырезаны
из первичного транскрипта и должен
произойти сплайсинг кодирующих
последовательностей. Оказалось, однако,
что процессинг - сложный многоступенчатый
процесс. Мутации в интроне гена bох-3
нарушают процессинг, потому что этот
интрон действительно транслируется,
чтобы мог образоваться белок, необходимый
для процессинга транскрипта гена box.
Последовательность в этом интронном
участке, определяющем синтез данного
белка, известна, так как этот участок
гена box был секвенирован.
Первый
этап сплайсинга первичного транскрипта
гена box, схематически представленного
на рисунке, приводит к образованию РНК,
содержащей кодирующую последовательность
bох-4/5 для цитохрома b, сцепленную с
маленьким кодирующим участком у конца
интрона box-3. Эта РНК, содержащая кодирующие
последовательности для цитохрома b и
интронные последовательности,
функционирует как мРНК, обеспечивая
синтез белка матуразы, необходимого
для следующей ступени процессинга. На
этом этапе происходит удаление
последовательности интрона box-3, с тем
чтобы образовалась мРНК, содержащая
только последовательности, кодирующие
цитохром b. Подобным же образом интрон
box-7 также, по-видимому, продуцирует
сходную, хотя и другую матуразу, поскольку
мутации box-3 и box-7 комплементарны друг
другу. Гипотетическая матураза box-7,
по-видимому, не только участвует в
процессинге гена цитохрома b, но необходима
также для процессинга транскрипта гена
oxi-3, поскольку мутации в интроне bох-7
воздействуют на синтез цитохромоксидазы.
В
этом гене имеется шесть кодирующих
последовательностей и пять интронов;
у него обнаружены три различных класса
мутаций. Мутации в кодирующих
последовательностях, как и следовало
ожидать, влияют на строение белка, и все
они входят в одну группу комплементации.
Мутации двух других классов необычны.
Три кластера локализованы в интронах.
Они в свою очередь образуют три различные
группы комплементации и блокируют
процессинг транскрипта гена цитохрома
b. Они оказывают также воздействие на
экспрессию гена oxi-3 - еще одного
разорванного митохондриального гена,
который кодирует субъединицу
1 цитохромоксидазы.
Мутации третьего класса локализованы
на границах между интронами и кодирующими
последовательностями.
Для того
чтобы могла образоваться функциональная
мРНК для цитохрома b, интронные
последовательности должны быть вырезаны
из первичного транскрипта и должен
произойти сплайсинг кодирующих
последовательностей. Оказалось, однако,
что процессинг - сложный многоступенчатый
процесс. Мутации в интроне гена bох-3
нарушают процессинг, потому что этот
интрон действительно транслируется,
чтобы мог образоваться белок, необходимый
для процессинга транскрипта гена box.
Последовательность в этом интронном
участке, определяющем синтез данного
белка, известна, так как этот участок
гена box был секвенирован.
Первый
этап сплайсинга первичного транскрипта
гена box, схематически представленного
на рисунке, приводит к образованию РНК,
содержащей кодирующую последовательность
bох-4/5 для цитохрома b, сцепленную с
маленьким кодирующим участком у конца
интрона box-3. Эта РНК, содержащая кодирующие
последовательности для цитохрома b и
интронные последовательности,
функционирует как мРНК, обеспечивая
синтез белка матуразы, необходимого
для следующей ступени процессинга. На
этом этапе происходит удаление
последовательности интрона box-3, с тем
чтобы образовалась мРНК, содержащая
только последовательности, кодирующие
цитохром b. Подобным же образом интрон
box-7 также, по-видимому, продуцирует
сходную, хотя и другую матуразу, поскольку
мутации box-3 и box-7 комплементарны друг
другу. Гипотетическая матураза box-7,
по-видимому, не только участвует в
процессинге гена цитохрома b, но необходима
также для процессинга транскрипта гена
oxi-3, поскольку мутации в интроне bох-7
воздействуют на синтез цитохромоксидазы.
 Роль
процессинга в регуляции генной экспрессии
выявляется также в примерах с α-амилазами из
печени и слюнной железы мышей. Хагенбюхль
и др.
обнаружили, что последовательности
мРНК в ферментах печени и ферменте
слюнной железы идентичны по своим
кодирующим участкам и нетранслируемым
3'-участкам. Однако нетранслируемые
5'-участки этих мРНК различны. Данные
названных авторов позволяют предполагать,
что эти белки кодируются одним и тем же
геном, но что его экспрессия, возможно,
регулируется тканеспецифичным
процессингом транскриптов. Очевидно,
что разорванные гены играют важную роль
в регуляции координированной генной
экспрессии у эукариот и что существование
подобной организации генов делает
возможной значительную эволюционную
пластичность.
Роль
процессинга в регуляции генной экспрессии
выявляется также в примерах с α-амилазами из
печени и слюнной железы мышей. Хагенбюхль
и др.
обнаружили, что последовательности
мРНК в ферментах печени и ферменте
слюнной железы идентичны по своим
кодирующим участкам и нетранслируемым
3'-участкам. Однако нетранслируемые
5'-участки этих мРНК различны. Данные
названных авторов позволяют предполагать,
что эти белки кодируются одним и тем же
геном, но что его экспрессия, возможно,
регулируется тканеспецифичным
процессингом транскриптов. Очевидно,
что разорванные гены играют важную роль
в регуляции координированной генной
экспрессии у эукариот и что существование
подобной организации генов делает
возможной значительную эволюционную
пластичность.
4