

Векторы на основе бактериофага λ
Плазмидные векторы позволяют клонировать фрагменты ДНК, размеры которых не превышают 10 т.п.н. Однако для решения задачи клонирования хромосомной ДНК даже небольшого организма, например – бактерии, необходимо создавать полные коллекции фрагментов этой ДНК, поэтому часто приходится работать с более крупными фрагментами. Для этого были разработаны векторы на основе бактериофага λ Ε. coli.
При проникновении фага λ в клетки E.coli. существуют два альтернативных пути развития событий:
1. Литический цикл – фаг начинает активно размножаться и примерно через 20 минут клетка разрушается с высвобождением до 100 новых фаговых частиц.
2. Состояние лизогении – фаговая ДНК включается в хромосому E.coli как профаг и реплицируется в клетке вместе с нормальными бактериальными клетками. Однако при неблагоприятных условиях (нехватка питания) запускается литический цикл (рис. 23):
1. При репликации кольцевой ДНК бактериофага λ образуется линейная молекула, состоящая из повторяющихся сегментов длиной примерно 50 т.п.н. Каждый из этих сегментов представляет собой полноразмерную фаговую ДНК, фланкированную липкими cos-сайтами - одноцепочечными 5'-«хвостами» из 12 нуклеотидов. Их называют липкими (cos) концами, поскольку они взаимно комплементарны и могут спариваться друг с другом подобно липким концам рестрикционных фрагментов.
2. Фаговая головка вмещает один такой сегмент, затем к головке присоединяется уже собранный отросток.
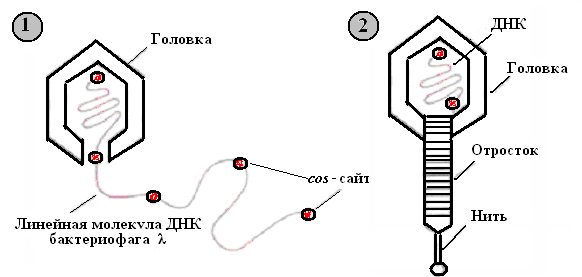
Рис. 23. Литический путь развития бактериофага λ
1 – упаковка в головку фага одного сегмента полноразмерной фаговой ДНК; 2 – сборка полноценной фаговой частицы.
Размер ДНК фага λ составляет примерно 50 т. п. н., причем значительная ее часть (около 20 т. п. н.) несущественна для размножения фага и отвечает за его встраивание в хозяйскую ДНК. В связи с этим возникла идея, что ее можно заменить фрагментом другой ДНК эквивалентного размера. Образующаяся рекомбинантная молекула будет реплицироваться в клетке как ДНК «рекомбинантного" фага, “вставшего" на литический путь развития. Рекомбинантные молекулы упаковывают в головки бактериофага λ in vitro и после добавления отростков получают инфекционные фаговые частицы (рис. 24).
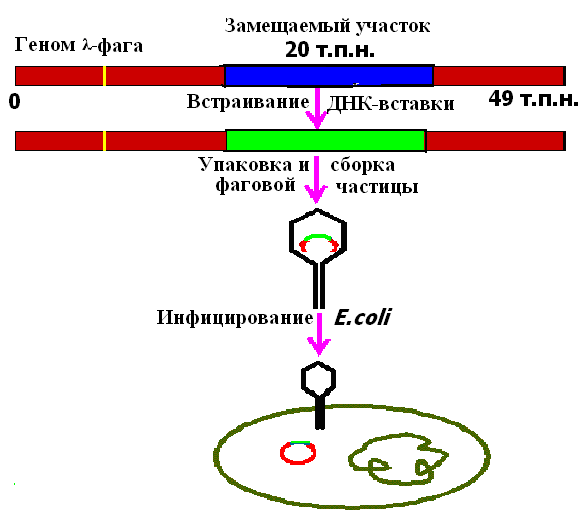
Рис. 24. Использование векторов на основе фага λ для клонирование фрагментов ДГК в клетках Ε. сoli.
Приготовление экстрактов для осуществления упаковки in vitro ДНК фага λ проводят с помощью двух штаммов E.coli, каждый из которых лизогенен в отношении определенного мутантного штамма фага λ (рис. 25). Один из мутантов не способен синтезировать белок А (один из полипептидов фаговой терминазы), другой – белок Е (белок головки фага). Оба этих белка необходимы для упаковки ДНК фага λ. “А”- и “Е”-экстракты смешивают и добавляют конкатемерную (сегменты полноразмерной фаговой ДНК, полимеризованные по cos-сайтам) ДНК фага, которая связывается с терминазой прежде, чем происходит разрезание в cos-сайтах, и упаковывается в фаговые головки.
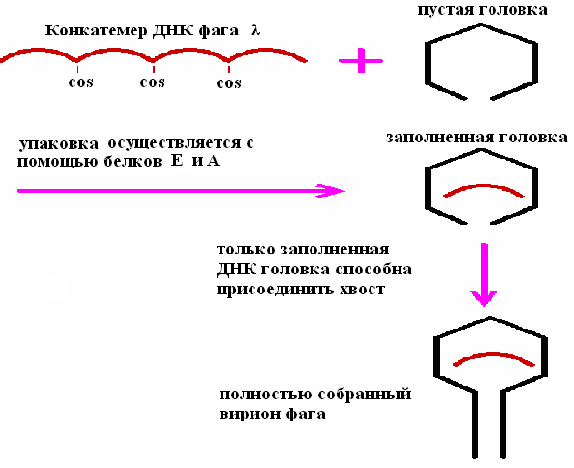
Рис. 25. Упаковка in vitro ДНК фага λ
При упаковке молекулы ДНК длиной менее 38 т.п.н. получается неинфекционная фаговая частица, а фрагменты длиной более 52 т, п. н. не умещаются в головку. Сегменты длиной 50 т. п. н. в линейной молекуле ДНК разделены cos-сайтами, и именно по этим сайтам разрезается молекула, когда очередной сегмент заполняет головку. Разрезание осуществляет фермент, находящийся у входа в головку.
Процесс введения рекомбинантной фаговой ДНК со встроенным фрагментом чужеродной генетической информации в клетки-реципиенты основан на естественном природном явлении – трансдукции фаговой ДНК.
Трансдукция (лат. transduction - перемещение) представляет собой процесс переноса бактериальной ДНК из одной клетки в другую бактериофагом. Таким образом, трансформация бактериальных клеток с помощью рекомбинантных ДНК на основе фаговой ДНК не требует специальной подготовки клеток-реципиентов или какого-либо специального приборного оснащения.
Для поиска клеток, содержащих фаги с рекомбинантными ДНК, используют методы молекулярной гибридизации и иммунологический скрининг, которые рассмотрим в следующем разделе.
Гибридные векторы
Фаговые векторы позволяют клонировать фрагменты ДНК длиной 15-25 т.п.н., что недостаточно для клонирования генов животных и растений, длина которых превышает 35-40 т.п.н. Требуемой емкостью обладают векторные молекулы, называемые космидами.
Космиды представляют собой небольшие плазмиды, в которые in vitro введены cos-сайты ДНК фага λ. Отсюда происходит название всего типа данных векторов (cosmid).
Космиды – один из видов гибридных векторов, которые реплицируются, используя плазмидный тип репликации, и обладают способностью упаковываться in vitro в оболочки частиц фага λ. Такие векторы могут включать до 40 т.п.н. чужеродной ДНК (рис. 26)
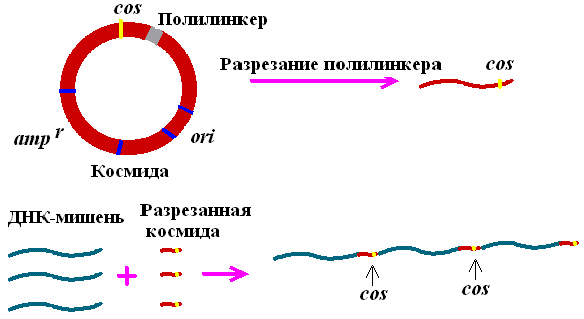
Рис. 26. Схема, демонстрирующая принцип конструирования рекомбинантных ДНК на основе космидного вектора
Наличие cos-сайтов в ДНК является единственным необходимым условием упаковываемости ДНК в фаговые частицы. Это означает, что последовательность нуклеотидов λ-ДНК, расположенная между двумя cos-сайтами, которая заключает в себе весь фаговый геном (35-45 т.п.н.), может быть замещена in vitro на аналогичный по длине фрагмент чужеродной ДНК и упакована в фаговые частицы. Такая искусственная фаговая частица оказывается нежизнеспособной. Однако, после адсорбции химерной фаговой частицы на поверхности бактериальной клетки, заключенная в ней ДНК проникает (вводится фаговой частицей) внутрь бактерии и начинает автономно реплицироваться как плазмида, размер которой составляет 30-40 т.п.н. Поскольку такая плазмида (космида) содержит в своем составе селектируемые маркеры в виде генов устойчивости к антибиотикам, ее поддерживают в бактериальных клетках путем выращивания бактерий на среде с соответствующими антибиотиками. Несмотря на то, что емкость космидных векторов значительно выше фаговых, эффективность клонирования в космидах ниже, хотя и достигает в ряде случаев 105-106 колоний на 1 мкг клонируемой ДНК
Стадия упаковки ДНК космид в фаговые частицы используется лишь для облегчения процесса введения рекомбинантных ДНК большого размера внутрь бактериальных клеток. Такой процесс имитирует проникновение фаговой хромосомы в бактерии во время фаговой инфекции.
В случае космид сходство между их проникновением в бактериальные клетки и фаговой инфекцией на этом заканчивается. Однако сходство является более глубоким в случае векторов, называемых фазмидами. Фазмиды представляют собой векторные молекулы ДНК, которые содержат в себе генетические элементы плазмид и хромосом бактериофагов. Они могут обладать емкостью в отношении клонируемой ДНК, характерной для лямбда-векторов, и существовать в определенных условиях в бактериальных клетках в виде плазмиды или же упаковываться в фаговые частицы in vivo при изменении этих условий.