

6. Секреторные ткани (наружные и внутренние): классификация, локализация в теле растения.
экзогенные и эндогенные
Одноклеточные и многоклеточные
Железистые волоски, нектарники, гидатоды, кристаллоносные клетки, слизевые клетки, масляные клетки, клетки с другими веществами, млечники, схизогенные, смоляные и камеденосные ходы, лизигенные полости.
Секреторные ткани распределены диффузно, имеют разное происхождение и большое морфологическое разнообразие. Экзогенные структуры- это производные протодермы, эндогенные- производные основной меристемы, прокамбия, камбия или специальных постоянных тканей: например флоэмы.
Могут выделять: углеводы, белки, гормоны, терпеноиды, алкалоиды, флавоноиды, дубильные вещества, капельно- жидкую воду.
7. Запасающие ткани: локализация, формы запасания угеводов (пластиды, оболочки клеток), беков, жиров.
Запасающая ткань — паренхима (основная ткань), выполняющая функцию хранения и запаса питательных веществ и воды.
Локализация: у семенных растений в эндосперме или зародыше семян. Многолетние растения, кроме того, накапливают запасные вещества в клубнях, луковицах, утолщенных корнях, сердцевине стеблей. Местом хранения резервных веществ может быть также паренхима проводящих тканей.
Запасающая ткань может превращаться в хлоренхиму. Лейкопла́сты образуются в запасающих тканях (клубнях, корневищах). Синтезируют и накапливают крахмал (амилопласты), жиры, белки. Лейкопласты содержат ферменты, с помощью которых из глюкозы, образованной в процессе фотосинтеза, синтезируется крахмал. На свету лейкопласты превращаются в хлоропласты.
Ф-ции: В запасающих тканях откладываются избыточные в данный период развития растения продукты метаболизма. Запасы хранятся в течение длительного срока (зимние запасы крахмала) либо потребляются и вновь пополняются в период вегетации (запасы воды). Расходование растением запасных веществ осуществляется гидролизом (при участии ферментов), приводящим к образованию растворимых в воде веществ, перемещающихся в места использования - в распускающиеся почки, в развивающийся проросток, в созревающие плоды и семена. Запасные вещества расходуются растением на рост, развитие, воспроизведение (при образовании нектара, мякоти плодов) и на дыхание, освобождающее рабочую энергию.
Строение: Среди тканей, запасающих пластические вещества, различают два типа: 1) ткани, накопляющие запасы в полостях клеток, и 2) ткани с запасами в полостях клеток и в их оболочках.
В тканях первого типа оболочки обычно тонки и снабжены мелкими простыми порами. Клетки содержат запасные вещества (чаще всего - сахара, крахмал, инулин, аминокислоты, белки):
а) в растворе (в корнеплоде свеклы, в сочных чешуях луковицы лука);
б) частью в растворе (амиды, белковые вещества), частью в твердом виде (крахмал, кристаллиды белка - в клубнях картофеля);
в) в твердом (алейроновые зерна) и полужидком (капли жирных масел) виде (в эндосперме клещевины);
г) почти полностью в твердом виде (в богатых алейроновыми зернами и крахмалом семядолях фасоли, гороха, чечевицы).
Запасающие ткани второго рода содержат обычно в клеточных полостях алейроновые зерна и жирные масла, а в оболочках клеток - отложения гемицеллюлоз либо амилоидов, а иногда слизей. Оболочки клеток сильно утолщаются и снабжаются многочисленными крупными простыми порами. Замыкающие пленки пор, а иногда и утолщенные участки клеточных стенок пронизываются плазмодесменными канальцами. Запасающие ткани этого рода содержатся в эндосперме семян кофе, финиковой пальмы, в семядолях люпинов, недотрог.
Ткани, запасающие воду (водоносные), состоят либо из живых паренхимных клеток с крупной центральной вакуолью с тонкими целлюлозными иногда ослизняющимися стенками(1), либо из мертвых трахеидоподобных клеток, отличающихся от обычных трахеид крупными размерами и изодиаметрической формой(2). Водозадерживающая ткань позволяет растениям выдерживать жару, а также недостаток воды в почве. При подсыхании листа вода из клеток водоносной ткани передается другим тканям, главным образом ассимиляционным. Стенки водоносных клеток сокращаются; когда лист снова обогащается водой, клетки принимают прежнюю форму.
|
Крахмальные зерна в клубне картофеля: 1 - простое крахмальное зерно, 2 - сложное, 3 - полусложное. |
|
1 — белковые кристаллы; 2 — аморфный белок матрикса алейронового зерна; 3 — глобоиды; 4 — липидные капли |

В запасающих тканях в особом типе лейкопластов - амилопластах часть сахаров откладывается в виде зерен вторичного крахмала. Рост крахмальных зерен происходит путем наложения новых слоев крахмала на старые, поэтому они имеют слоистую структуру. Если имеется один центр- это простое зерно, если более, то образуется сложное зерно. Полусложное зерно формируется в тех случаях, когда крахмал сначала откладывается вокруг нескольких точек, а затем после соприкосновения простых зерен вокруг них возникают общие слои.
Запасные белки наиболее часто встречаются в виде алейроновых зерен в клетках семян бобовых, гречишных, злаков. Алейроновые зерна образуются при созревании семян из высыхающих вакуолей. Алейроновое зерно окружено тонопластом и содержит белковый матрикс, в который погружены белковый кристалл ромбоэдрической формы и глобоид фитина (запасной фосфор). Это сложное алейроновое зерно (у льна, тыквы, подсолнечника). Алейроновые зерна, содержащие только аморфный белок, называют простыми (у бобовых, риса, кукурузы, гречихи). При прорастании семян алейроновые зерна набухают, белки и фитин подвергаются ферментативному расщеплению, продукты которого используются растущим зародышем. При этом алейроновые зерна постепенно превращаются в типичные вакуоли, лишенные белка.
Широко распространено у растений отложение запасных жиров в виде липидных капель в цитоплазме. Наиболее богаты ими семена и плоды. Во время прорастания семян они гидролизуются с образованием растворимых углеводов.
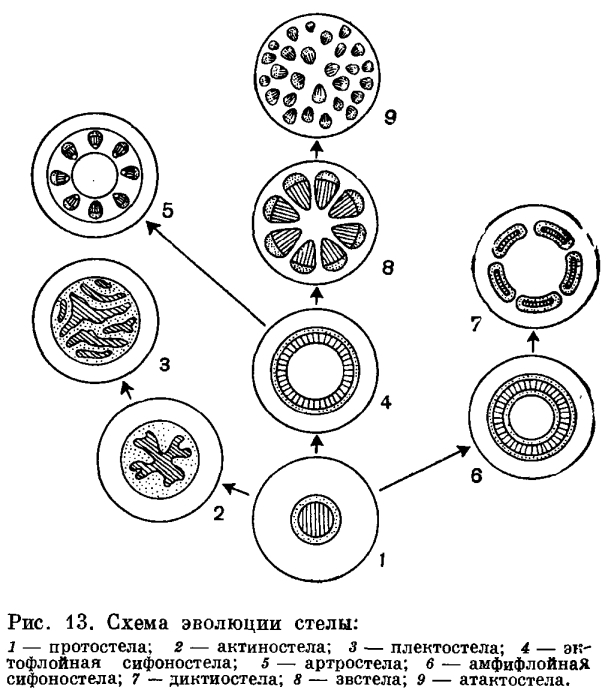
Билет №8. Стела: строение, типы
Стела- первичную проводящую систему осевых органов высш раст вместе со связанными с ней паренхимными тканями.
протостела- примитивный тип, проводящая ткань образует сплошную массу и центральное ядро из ксилемы окружено слоем флоэмы. Это исходный тип, от которого произошли все другие. Была характерна для риниофитов и других вымерших форм. В современной флоре- плаун, нек папоротники и в корнях почти всех растений. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы =гаплостела. Более подвинутым типом- протостела с радиальными выступами ксилемы = актиностела- была характерна для девонского рода астероксилон, встречается у некоторых примитивных видов плаунов.
Плектостела (плетеный)- дальнейшая специализации актиностелы, у стеблей большинства видов плауна. ксилема разбита на пластинки, расположенные параллельно или радиально.
Сифоностела (труба)- из протостелы. трубчатое строение, обладает центральной паренхимной частью(сердцевиной). Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в проводящем цилиндре для непрерывной связи коры и сердцевины. характерны для папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей получается рассеченная диктиостела (сеть), у папоротников.
Полициклическая сифоностела- наиболее сложный тип- у нек папоротников. Имеют внутреннюю проводящую систему, связанную с внешней сифоностелой. Эта внутренняя связь бывает всегда у узла. два или больше концентрических цилиндров проводящей ткани. В некоторых случаях внешний цилиндр является сифоностелой, а в других — диктиостелой.
артростела (член, сустав)- у хвощей, из сифоностелы. членистое строение и наличие центральной полости и протоксилемных лакун (каринальных каналов).
эвстела (хорошо)- при рассечения сифоностелы, у большинства семенных растений.
атактостела (беспорядочный). у травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны.
Билет №9. Общая характеристика стебля.
Стебль- осевую часть побега, с листья, почки, цветы и плоды. функция- опорная, проводящая, запасающая + орган вегетативного размножения, орган фотосинтеза.
Стебель, как и корень, развивается из зародыша семени, радиальная симметрия, неограниченный рост. Но в отличие от корня обладает свойством - геотропизма и + фототропизма. У основной массы травянистых растений стебли живут в течение одного вегетационного периода.
2 типа: Деревянистый –многолетний, утолщающийся, образованный одревесневающими тканями (береза, виноград, смородина). Травянистый – обычно существующий 1 вегетационный период, не утолщающийся (слабо) и неодревесневающий (слабо) (крапива, лебеда).
Ф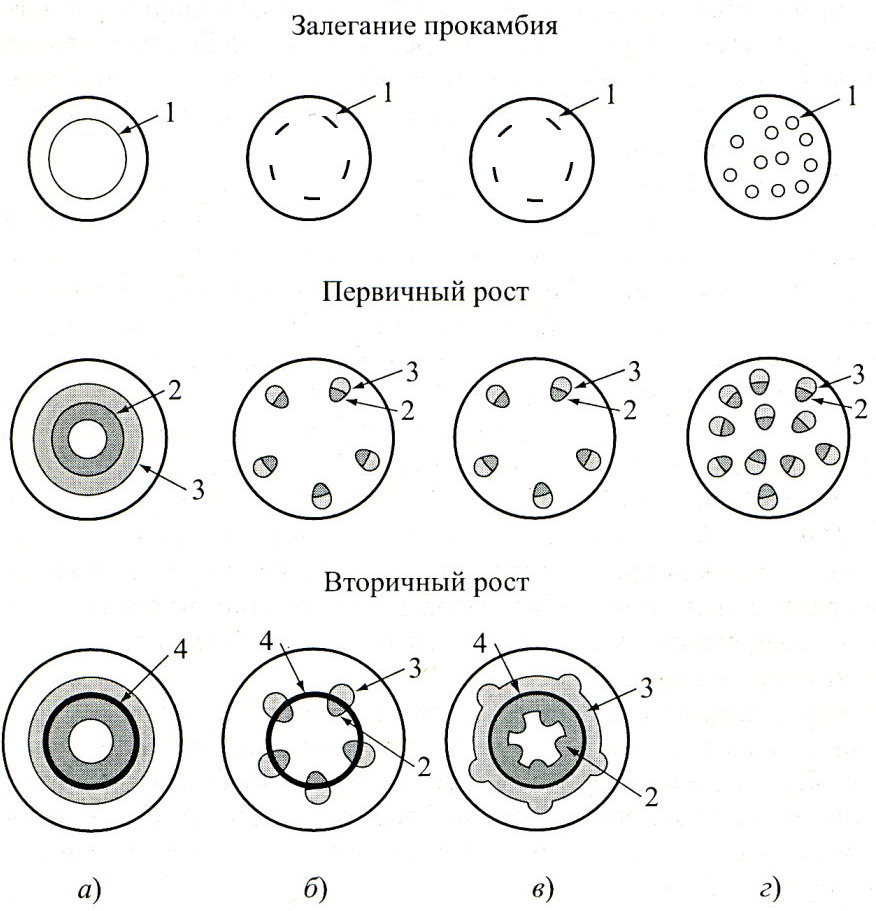 ормирование
различных типов стеблей: а – сифоностела;
б – эвстела, пучковое строение, проводящие
пучки расположены кольцом; в – эвстела,
пучковое строение; при вторичном росте
формируется переходное строение; г –
атактостела, пучковое строение, проводящие
пучки расположены беспорядочно,
вторичного роста нет. 1 – прокамбий; 2 –
ксилема; 3 – флоэма; 4 – камбий.
Анатомия. более
разнообразно, чем у корня. Первичное
строение
сохраняется всю жизнь у растений без
камбия. Снаружи стебель покрыт эпидермой,
под ней – первичная кора, окружающая
стелу. Прокамбий,
первичная
латеральная образовательная ткань,
образующийся из конуса нарастания,
полностью расходуется на формирование
первичной флоэмы и ксилемы. Пучки
закрытые.У голосеменных и двудольных
часть прокамбия преобразуется в камбий.
Вторичное
строение
формируется за счет деятельности
вторичной боковой меристемы – камбия,
формирующего вторичную ксилему и
вторичную флоэму.
Стела
эволюционировала от протостелы риниофитов
(в центре Кс, на периферии Ф) до атактостелы
однодольных. Затем сформировалась
сифоностела с сердцевиной в центре (у
папоротников), эвстела у большинства
двудольных растений и атактостела
однодольных.
ормирование
различных типов стеблей: а – сифоностела;
б – эвстела, пучковое строение, проводящие
пучки расположены кольцом; в – эвстела,
пучковое строение; при вторичном росте
формируется переходное строение; г –
атактостела, пучковое строение, проводящие
пучки расположены беспорядочно,
вторичного роста нет. 1 – прокамбий; 2 –
ксилема; 3 – флоэма; 4 – камбий.
Анатомия. более
разнообразно, чем у корня. Первичное
строение
сохраняется всю жизнь у растений без
камбия. Снаружи стебель покрыт эпидермой,
под ней – первичная кора, окружающая
стелу. Прокамбий,
первичная
латеральная образовательная ткань,
образующийся из конуса нарастания,
полностью расходуется на формирование
первичной флоэмы и ксилемы. Пучки
закрытые.У голосеменных и двудольных
часть прокамбия преобразуется в камбий.
Вторичное
строение
формируется за счет деятельности
вторичной боковой меристемы – камбия,
формирующего вторичную ксилему и
вторичную флоэму.
Стела
эволюционировала от протостелы риниофитов
(в центре Кс, на периферии Ф) до атактостелы
однодольных. Затем сформировалась
сифоностела с сердцевиной в центре (у
папоротников), эвстела у большинства
двудольных растений и атактостела
однодольных.
Рис. Стебля липы: 1 — эпидерма; 2 — пробка; 3 — первичная кора; 4 — луб(вт кора); 5 — сердцевинный луч; 6 — камбий; 7 — осенний сосуд; 8 — весенний сосуд; 9 — первичная древесина; 10 — сердцевина; 11 — чечевичка.
П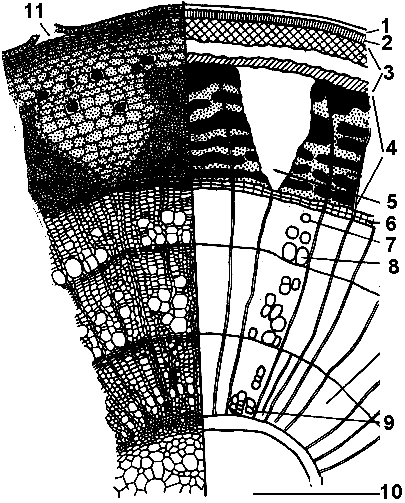 ервичное
строение.
ервичное
строение.
эпидерма, первичная кора (паренхима, часто с хлоропластами). Внутр слой первичной коры - эндодерма = крахмалоносное влагалище (много крахмальных зерен).
Наружный слой стелы= как и у корня, перицикл. Сохраняет меристематическую активность –могут закладываться придаточные почки и придаточные корни. в центре обр-ся сердцевина из паренхимных кл.
Вторичное строение. прокамбий закладывается в виде отдельных групп клеток. После образования камбия закладывается вторичная Кс и Ф- открытые пучки, Если межпучковый камбий образует клетки паренхимы, то образуется эвстела, если ксилему и флоэму, то образуется переходный тип строения стебля, при котором проводящие ткани образуют кольцо неправильной формы. Для древесных растений - непучковый тип строения- прокамбий образуется сплошным кольцом, формируя первичную Кс и Ф, а затем образуется камбий и вторичный рост стебля. Под эпидермой- пробковый камбий – феллоген (откладывает наружу пробки, внутрь –феллодерму). Пробка, феллоген и феллодерма образуют вторичный покров – перидерму. Под нек устьицами закладываются чечевички. Под перидермой- кора (первичная и вторичная), камбий, древесина и сердцевина. Под первичной корой- Ф (луб) – вторичная кора с проводящими тканями – ситовидные клетки и ситовидные трубки с клетками спутницами, механические ткани – лубяные волокна и основные ткани – клетки лубяной паренхимы, выполняющие запасающую функцию. Во вторичной коре хорошо просматриваются сердцевинные лучи. На срезах сердцевинные лучи имеют вид светлых треугольников. Они чередуются с трапецевидными участками флоэмы. Под корой находится камбий, вторичная латеральная меристема. Большая часть стебля образована клетками, возникшими в результате деятельности камбия, располагающегося между вторичной корой и древесиной. Именно благодаря ему происходят вторичные изменения в строении стебля. Обычно в древесину камбий откладывает большее число производных, чем наружу, соотношение 4:1 соответственно. Весной клетки камбия активно делятся, с приближением осени деятельность камбия ослабевает, и зимой он вступает в период покоя.
Древесина. Внутрь от камбия откладываются клетки древесины (вторичной ксилемы), в состав которой входят сосуды, трахеиды, древесная паренхима и древесная склеренхима (волокна). Особенностью ксилемы является одревеснение клеточных стенок (за исключением клеток древесной паренхимы). Вторичная ксилема составляет основную массу (9/10 объема) древесного стебля.
Годичные кольца – прирост древесины за 1 вегетационный период. Весной камбий откладывает широкопросветные и тонкостенные сосуды и трахеиды, к которым примыкают клетки древесной паренхимы. Осенью в древесине преобладают узкопросветные и толстостенные сосуды, трахеиды и древесные волокна. Переход от весенней древесины к осенней постепенный, наоборот- резкий. Весенняя древесина светлее.
Сердцевина. В центре стебля, образованная округлыми паренхимными клетками. окружена сосудами первичной ксилемы. Горизонтальный транспорт органических веществ в клетки сердцевины и обратно осуществляется по сердцевинным лучам.