

1)Ткани растения – определение. Классификация тканей. Покровные ткани: эпидерма, экзодерма, корка, перидерма (локализация, происхождение, состав)
Ткани – комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций. возникли у растений в связи с переходом водорослей к наземному образу жизни (сопровождался дифференциацией однородного слоевища водных обитателей на основные вегетативные органы: корень и побег) классифицируют в зависимости от выполняемой функции.
сложные - состоящие из разнородных элементов, выполняющих разные функции, и простые - состоящие из однородных элементов. 1) образовательные, меристемы; 2)основные; 3) покровные; 4) выделительные; 5) механические; 6) проводящие. Последние 5 типов объединяют в постоянные ткани. Они образуются из меристем путем дифференциации клеток.
Покровные ткани
на поверхности органов растений на границе с внешней средой.
Клетки плотно сомкнуты, защищают внутренние части растения от неблагоприятных внешних воздействий, излишнего испарения и иссушения, резкой перемены температуры, проникновения микроорганизмов, служат для газообмена и транспирации.
В соответствии с происхождением из различных меристем выделяют первичные и вторичные покровные ткани. Ризодерма (эпиблема) – первичная однослойная поверхностная ткань корня. Образуется из протодермы – наружного слоя клеток апикальной меристемы корня.
функция– всасывание, избирательное поглощение из почвы воды с мин в-вами. выделение веществ, действующих на субстрат и преобразующих его.
Клетки тонкостенные, с вязкой цитоплазмой и большим количеством митохондрий.
Характерной особенностью является образование у части клеток корневых волосков. Эпидерма - первичная покровная ткань, образующаяся из протодермы конуса нарастания побега. покрывает листья, стебли, цветки, плоды и семена.
ф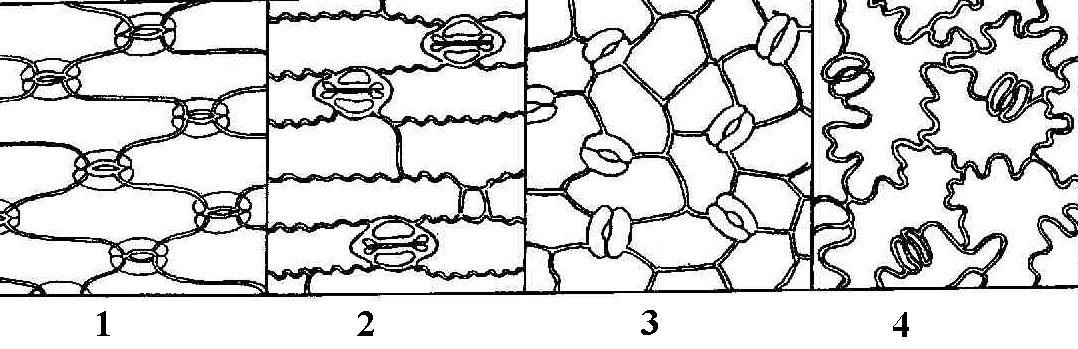 ункция–
регуляция газообмена и транспирации.
препятствует проникновению внутрь
растения болезнетворных организмов,
защищает внутренние ткани от механических
повреждений и придает органам прочность.
Через эпидерму могут выделяться наружу
эфирные масла, вода, соли. может
функционировать как всасывающая ткань.
принимает участие в синтезе различных
веществ, в восприятии раздражений, в
движении листьев.
Эпидерма
- сложная ткань, в ее состав входят
морфологически различные типы клеток:
1) основные клетки эпидермы;
2) замыкающие и побочные клетки
устьиц; 3) трихомы.
Основные
клетки эпидермы –
живые клетки таблитчатой формы. плотно
сомкнуты, межклетники отсутствуют.
Боковые стенки часто извилистые, что
повышает прочность их сцепления, реже
прямые. Наружные стенки толще. Их
внутренний, более мощный, слой состоит
из целлюлозы и пектиновых веществ;
наружный слой подвергается кутинизации.
Рис.
Эпидерма листа (с поверхности): 1 -
ирис; 2 - кукуруза; 3 – арбуз; 4 -
буквица.
Устьица –
образования для регуляции транспирации
и газообмена. состоит из
2 замыкающих клеток бобовидной
формы, между которыми находится устьичная щель,
которая может расширяться и сужаться.
ункция–
регуляция газообмена и транспирации.
препятствует проникновению внутрь
растения болезнетворных организмов,
защищает внутренние ткани от механических
повреждений и придает органам прочность.
Через эпидерму могут выделяться наружу
эфирные масла, вода, соли. может
функционировать как всасывающая ткань.
принимает участие в синтезе различных
веществ, в восприятии раздражений, в
движении листьев.
Эпидерма
- сложная ткань, в ее состав входят
морфологически различные типы клеток:
1) основные клетки эпидермы;
2) замыкающие и побочные клетки
устьиц; 3) трихомы.
Основные
клетки эпидермы –
живые клетки таблитчатой формы. плотно
сомкнуты, межклетники отсутствуют.
Боковые стенки часто извилистые, что
повышает прочность их сцепления, реже
прямые. Наружные стенки толще. Их
внутренний, более мощный, слой состоит
из целлюлозы и пектиновых веществ;
наружный слой подвергается кутинизации.
Рис.
Эпидерма листа (с поверхности): 1 -
ирис; 2 - кукуруза; 3 – арбуз; 4 -
буквица.
Устьица –
образования для регуляции транспирации
и газообмена. состоит из
2 замыкающих клеток бобовидной
формы, между которыми находится устьичная щель,
которая может расширяться и сужаться.
Т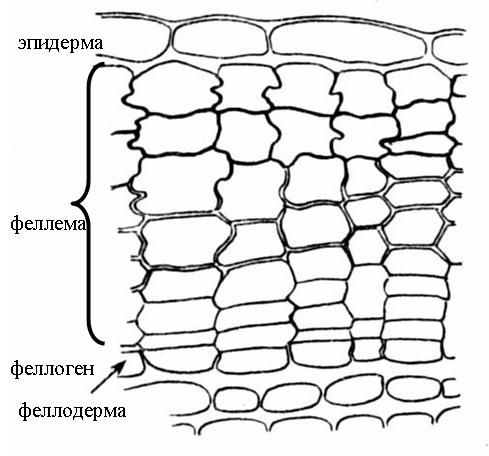
 рихомы
(наружные
одно- или многоклеточные выросты):
1) кроющие, 2) железистые. Железистые образуют
вещества, которые рассматриваются как
выделения. Кроющие имеют
вид простых, разветвленных или звездчатых
волосков, могут длительное время
оставаться живыми, но чаще они быстро
отмирают и заполняются воздухом.
Густой
слой волосков отражает часть солнечных
лучей и уменьшает нагрев, что снижает
транспирацию.
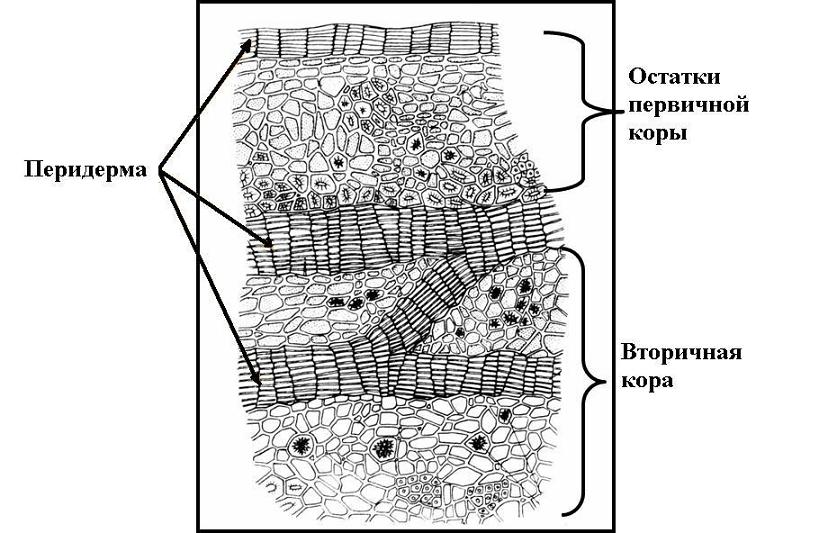
К вторичным покровным
тканям относятся: 1) перидерма и
2) корка, или ритидом.
Перидерма –
сложная многослойная покровная ткань,
которая приходит на смену первичным
покровным тканям – ризодерме и эпидерме.
покрывает корни вторичного строения и
стебли многолетних побегов. может
возникнуть и в результате залечивания
поврежденных тканей раневой меристемой.
из
3 комплексов клеток:
1) феллема(пробка)-
защитные функции; 2) феллоген(пробковый камбий)-
образует перидерму в целом; 3) феллодерма
(пробковая паренхима)- питает
феллоген.
Феллема (пробка)- несколько
плотных слоев таблитчатых мертвых
клеток, без межклетников. Вторичные
клеточные стенки состоят из чередующихся
слоев суберина и воска, что делает их
непроницаемыми для воды и газов. не
имеют протопласта и заполнены воздухом.
функция–
защита от потери влаги, предохраняет
от проникновения болезнетворных
организмов, дает механическую защиту
стволам и ветвям.
Феллоген
(пробковый
камбий) – вторичная латеральная
меристема. 1 слой меристематических
клеток, откладывающих клетки пробки
наружу и клетки феллодермы внутрь.
залечивает нанесенные повреждения,
образуя новые слои пробки.
рихомы
(наружные
одно- или многоклеточные выросты):
1) кроющие, 2) железистые. Железистые образуют
вещества, которые рассматриваются как
выделения. Кроющие имеют
вид простых, разветвленных или звездчатых
волосков, могут длительное время
оставаться живыми, но чаще они быстро
отмирают и заполняются воздухом.
Густой
слой волосков отражает часть солнечных
лучей и уменьшает нагрев, что снижает
транспирацию.
К вторичным покровным
тканям относятся: 1) перидерма и
2) корка, или ритидом.
Перидерма –
сложная многослойная покровная ткань,
которая приходит на смену первичным
покровным тканям – ризодерме и эпидерме.
покрывает корни вторичного строения и
стебли многолетних побегов. может
возникнуть и в результате залечивания
поврежденных тканей раневой меристемой.
из
3 комплексов клеток:
1) феллема(пробка)-
защитные функции; 2) феллоген(пробковый камбий)-
образует перидерму в целом; 3) феллодерма
(пробковая паренхима)- питает
феллоген.
Феллема (пробка)- несколько
плотных слоев таблитчатых мертвых
клеток, без межклетников. Вторичные
клеточные стенки состоят из чередующихся
слоев суберина и воска, что делает их
непроницаемыми для воды и газов. не
имеют протопласта и заполнены воздухом.
функция–
защита от потери влаги, предохраняет
от проникновения болезнетворных
организмов, дает механическую защиту
стволам и ветвям.
Феллоген
(пробковый
камбий) – вторичная латеральная
меристема. 1 слой меристематических
клеток, откладывающих клетки пробки
наружу и клетки феллодермы внутрь.
залечивает нанесенные повреждения,
образуя новые слои пробки.
Феллодерма (пробковая паренхима) - основная ткань из живых паренхимных клеток. Газообмен и транспирация в органах, покрытых перидермой, происходят через чечевички. У большинства древесных растений на смену гладкой перидерме приходит трещиноватая корка (ритидом).Корка возникает в результате многократного заложения новых прослоек перидермы во все более глубоких слоях коры. Живые клетки между прослойками погибают. Т.о., корка состоит из чередующихся слоев пробки и прочих отмерших тканей коры. 2) Фотосинтезирующая ткань: локализация в теле растения, характер мезофилла, строение хлоропластов, сезонная динамика пластид.
Ассимиляционные ткани– фотосинтез. по форме клеток относятся к паренхимным: у высших растений имеют зеленую окраску (хлоренхима). Имеют тонкие целлюлозные стенки. У немногих растений стенки клеток столбчатой хлоренхимы снабжены внутренними утолщениями. Протопласт состоит из постенного слоя протоплазмы(ядро и хлоропласты в один слой), центральная часть клетки занята крупной вакуолью.
Хлоренхима -непосредственно под кожицей листьев и стебле. К хлоренхиме обычно примыкает кожица с устьицами, через которые подается с воздухом необходимый для работы фотосинтеза углекислый газ.
Хлоренхима(мезофилл) дифференцирована на столбчатую и губчатую ткань. Столбчатая- удлиненные клетки цилиндрической формы, межклетники расположены между продольно вытянутыми частями оболочек соседних клеток. Губчатая- округленные кл. Межклетники образуют сложную сетчатую систему, с многочисленными разветвлениями.
Эмбриональные клетки содержат бесцветные пропластиды. В зависимости от типа ткани они развиваются в хлоропласты; хромопласты; лейкопласты.
Хлоропласты — находятся в паренхимных клетках зеленых частей высших растений. Снаружи покрыт оболочкой из 2 липопротеиновых мембран. Внутренняя мембрана образует складчатые впячивания внутрь матрикса(стромы). В зрелом хлоропласте видны 2 типа внутренних мембран. Это- мембраны, образующие плоские, протяженные ламеллы, и мембраны тилакоидов, плоских дисковидных вакуолей. ламеллы лежат параллельно и не образуют связей между собой. мембранные тилакоиды- плоские замкнутые мембранные мешки, в форме диска- образуют стопки(граны). Ламеллы стромы связывают между собой отдельные граны. Но полости камер тилакоидов замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. В матриксе- ДНК, рибосомы; первичное отложение запасного полисахарида (крахмальные зерна). Пигменты: хлорофилл А (сине-зеленый) - 70 % (у высших растений и зеленых водорослей); хлорофилл В (желто-зеленый) - 30 % (там же); хлорофилл С, D и E встречается реже - у других групп водорослей; каротиноиды и антоцианы. При осеннем пожелтении листьев разрушение хлоропластов и образование хромопластов приводит к утилизации белков и азотсодержащих соединений, которые перед листопадом оттекают в другие органы растения.
Билет №3
Механические ткани- скелет, скрепляет ткани и части органов между собой. прочность, способность противостоять действию тяжести собственных органов, порывам ветра, дождю, вытаптыванию.
Клетки разнообразны по форме, но у всех сильно утолщенные клеточные стенки, даже после отмирания протопласта продолжают выполнять опорную функцию. Различают два типа механических тканей: 1) колленхиму и 2) склеренхиму. Колленхима – механическая ткань растущих органов, возникает очень рано.
Клетки. живые, вытянуты по оси органа, с тупыми или скошенными концами. часто есть хлоропласты. Утолщены неравномерно, не одревесневают. Граница между первичной и вторичной стенками не выражена, в утолщениях чередуются слои целлюлозы и сильно обводненные слои, богатые пектинами и гемицеллюлозами.
Ф
 ункции
опорной ткани колленхима может выполнять
только в состоянии тургора. Если растение
теряет воду, тонкие участки клеточных
стенок складываются «гармошкой», побеги
теряют упругость и обвисают. Колленхима
располагается сразу за покровной тканью.
В корнях не встречается. характерна для
двудольных.
3
типа колленхимы: 1) уголковую,
2) пластинчатую и
3) рыхлую.
Уголковая
колленхима-
стенки, утолщенные в углах клеток.
Утолщения соседних клеток смыкаются,
образуя трех – пятиугольники.
Часто
встречается в стеблях травянистых
растений, черешках листьев, вдоль главной
жилки листа.
ункции
опорной ткани колленхима может выполнять
только в состоянии тургора. Если растение
теряет воду, тонкие участки клеточных
стенок складываются «гармошкой», побеги
теряют упругость и обвисают. Колленхима
располагается сразу за покровной тканью.
В корнях не встречается. характерна для
двудольных.
3
типа колленхимы: 1) уголковую,
2) пластинчатую и
3) рыхлую.
Уголковая
колленхима-
стенки, утолщенные в углах клеток.
Утолщения соседних клеток смыкаются,
образуя трех – пятиугольники.
Часто
встречается в стеблях травянистых
растений, черешках листьев, вдоль главной
жилки листа.
П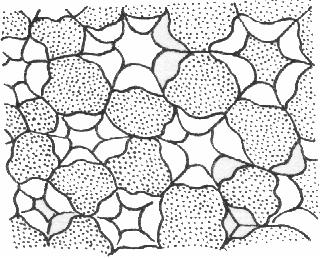 ластинчатая-
утолщения тангенциальных (параллельных
поверхности органа) стенок, которые
располагаются параллельными слоями,
радиальные стенки остаются тонкими.
Она встречается, чаще всего, в молодых
стеблях древесных растений.
ластинчатая-
утолщения тангенциальных (параллельных
поверхности органа) стенок, которые
располагаются параллельными слоями,
радиальные стенки остаются тонкими.
Она встречается, чаще всего, в молодых
стеблях древесных растений.
Рыхлая- хорошо выраженные межклетники. Утолщаются части стенок, прилегающ к межклетным пространствам. У нек травянистых растений (лопух, дурман). Сочетает признаки уголковой колленхимы и аэренхимы.
С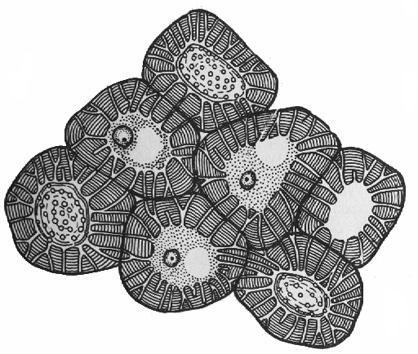 клеренхима-
во
всех органах: корнях, стеблях, листьях,
плодах, цветках, семенах. Равномерно
утолщенные
одревесневшие
стенки. Полость клетки мала, поры простые,
щелевидные, немногочисленные. Протопласт
рано отмирает, и опорную функцию выполняют
мертвые клетки. 2 типа склеренхимы:
1) волокна и
2) склереиды,
различающиеся формой.
Волокна–
прозенхимные кл, вытянутые в длину,
заостренные на концах. Прочность органов
на сжатие, растяжение и изгибы. Прочность
волокон повышается благодаря тому, что
фибриллы целлюлозы проходят в них
винтообразно, меняя направление во
внешних и внутренних витках.
Волокна
в коровой части (флоэма)- лубяные
(неодревесневающей клеточной стенкой).
Волокна в древесине (ксилеме)- древесинные.
стенки всегда одревесневшие, длина их
не превышает 2 мм.
клеренхима-
во
всех органах: корнях, стеблях, листьях,
плодах, цветках, семенах. Равномерно
утолщенные
одревесневшие
стенки. Полость клетки мала, поры простые,
щелевидные, немногочисленные. Протопласт
рано отмирает, и опорную функцию выполняют
мертвые клетки. 2 типа склеренхимы:
1) волокна и
2) склереиды,
различающиеся формой.
Волокна–
прозенхимные кл, вытянутые в длину,
заостренные на концах. Прочность органов
на сжатие, растяжение и изгибы. Прочность
волокон повышается благодаря тому, что
фибриллы целлюлозы проходят в них
винтообразно, меняя направление во
внешних и внутренних витках.
Волокна
в коровой части (флоэма)- лубяные
(неодревесневающей клеточной стенкой).
Волокна в древесине (ксилеме)- древесинные.
стенки всегда одревесневшие, длина их
не превышает 2 мм.
Склереиды – мертвые паренхимные кл с толстыми одревесневшими стенками, пронизанными поровыми каналами. Встречаются как поодиночке (идиобласт), так и группами. В зависимости от формы: каменистые клетки и астросклереиды.
Каменистые клетки- округлой формы. косточки вишни, сливы, персика, скорлупа грецкого ореха. в мякоти плодов груши, айвы, рябины, в корнях хрена среди тонкостенных клеток.
Астросклереиды имеют ветвистую форму с отростками, направленными в разные стороны. Они располагаются в виде идиобластов в мезофилле листьев некоторых растений (камелия, маслина, кубышка), скрепляя рыхлые ткани подобно шпильке в волосах. №4. Флоэма: функция, происхождение, закономерности организации, состав, варианты строения, локализация в теле растения.
Перемещения веществ в виде водных растворов, т.е. важно наличие тесных контактов между обеими Кс и Ф, объединение в систему взаимосвязанных проводящих пучков.: Кс(древесина) и флоэма(луб). По флоэме в нисходящем направлении движутся продукты ассимиляции.
Пучки. Коллатеральный - флоэма обращена к периферии, а ксилема - к центру (в стеблях и листьях многих семенных растений).
Биколлатеральный – Ф прилегает к Кс с обеих сторон, наружный участок Ф больше, чем внутренний (нек. 2-дольным, напр, тыквенным).
Концентрический – одна из тканей полностью окружает другую: ксилема вокруг флоэмы - амфивазальный пучок (у однодольных; у драцены); ксилема окружена флоэмой – амфикрибральный пучок (у папоротников).
Радиальный – чередование ксилемы и флоэмы (корневая зона).
Открытые пучки – между флоэмой и ксилемой есть камбий, пучок разрастается (2- дольные и голосеменные).
Закрытые - камбия нет, пучок не разрастается (однодольные; нек. 2-дольные, т.к. камбий у них рано перестает функционировать - лютик).
Флоэму и ксилему образуют специальные васкулярные меристемы – прокамбий и камбий. Проводящие ткани, возникшие из прокамбия, относят к первичным, а ткани, развивающиеся из производных камбия, - ко вторичным. Первичные ксилему и флоэму делят на прото-, мета- ксилему и флоэму. Первичные проводящие ткани характерны для всех высших растений, вторичные - для голосеменных и 2-дольных покрытосеменных.
Флоэма - сложная проводящая ткань, транспорт продуктов фотосинтеза от листьев к местам их использования или отложения (к конусам нарастания , подземным органам, зреющим семенам и плодам).
Первичная флоэма дифференцируется из прокамбия , вторичная флоэма (луб) - производная камбия . В стеблях флоэма находится обычно снаружи от ксилемы , а в листьях она обращена к нижней стороне пластинки. Первичная и вторичная флоэмы, помимо различной мощности ситовидных элементов, отличаются тем, что у первой отсутствуют сердцевинные лучи.
В состав флоэмы входят ситовидные элементы, паренхимные клетки, элементы сердцевинных лучей и механические элементы. Большинство клеток нормально функционирующей флоэмы живые. Отмирает лишь часть механических элементов. Собственно проводящую функцию осуществляют ситовидные элементы. Различают два их типа: ситовидные клетки и ситовидные трубки. Терминальные стенки ситовидных элементов содержат мелкие сквозные канальцы, собранные группами в ситовидные поля. У ситовидных клеток ситовидные поля располагаются на боковых стенках. Ситовидные клетки - основной проводящий элемент флоэмы у высших растений, исключая покрытосеменные . Клеток-спутниц у ситовидных клеток нет.
Ситовидные трубки покрытосеменных более совершенны. Они состоят из члеников, располагающихся один над другим. Длина члеников -150-300 мкм. Поперечник ситовидных трубок составляет 20-30 мкм. Эволюционно членики возникли из ситовидных клеток.
Ситовидные поля члеников находятся на концах и соседние образуют ситовидную пластинку. Членики формируются из вытянутых клеток прокамбия или камбия: материнская клетка меристемы делится в продольном направлении и производит две клетки: членик и клетку-спутницу + поперечное деление клетки-спутницы с последующим образованием двух-трех подобных клеток, расположенных продольно одна над другой. При своем формировании членик имеет постенную цитоплазму, ядро и вакуоль. С началом функциональной деятельности он заметно вытягивается. На поперечных стенках появляется множество мелких отверстий-перфораций, образующих канальцы диаметром несколько микрометров, через которые из членика в членик проходят цитоплазматические тяжи. На стенках канальцев откладывается полисахарид каллоза , сужающий их просвет, но не прерывающий цитоплазматические тяжи.
По мере развития членика ситовидной трубки в протопласте образуются слизевые тельца- содержимое сливается в единую массу, цитоплазма становится проницаемой для растворов орг и неорг веществ. Слизевые тельца сливаются, образуя слизевой тяж и скопления около ситовидных пластинок. На этом формирование членика ситовидной трубки завершается.
По мере старения ситовидные трубки закупориваются каллозой (образующей мозолистое тело) и отмирают. Отмершие ситовидные трубки обычно сплющиваются.
Паренхимные элементы флоэмы (лубяная паренхима) состоят из тонкостенных клеток. В них откладываются запасные пит в-ва и отчасти по ним осуществляется ближний транспорт ассимилянтов. У голосеменных клетки-спутницы отсутствуют и их роль выполняют прилегающие к ситовидным клеткам немногочисленные клетки лубяной паренхимы.
Сердцевинные лучи, продолжающиеся во вторичной флоэме, -из тонкостенных паренхимных клеток. Ближний транспорт ассимилянтов.