

Класифікація губок
Губок за будовою та складом їх скелету поділяють на
класи: 1. Вапнякові губки (С а 1 с і 8 р о п § і а);
Скляні губки (Н у а 1 о з р о п § і а); 3. Звичайні губки (Б е га о 8 р о п £ і а); 4. Археоціати (АгсЬае-
о с у а і Ь а).
У представників класу Вапнякові губки скелет складається зі спікул СаС03. Це виключно морські, невеликих розмірів мілководні поодинокі чи колоніальні губки. Є представники, побудовані по типу аскон, сикон, лейкон.
До класу Скляні губки належать морські, здебільшого глибоководні губки. Переважно поодинокі форми типу сикон, висотою до 50 см і більше. Мезохіл майже зовсім відсутній. Скелет побудований із голок 8Ю2. У губки монорафіс (МопогЬарЬіз сЬипі) циліндричне тіло, висота якого близько 1 м, пронизане голкою у вигляді вісі, що має довжину 3 м і встромляється в ґрунт.
До класу Звичайні губки належить більшість представників, що населяють солоні та прісні водойми. Це колоніальні або поодинокі форми, побудовані за лейкоїдним типом. Скелет утворений спікулами із 8Ю2, з’єднаними спонгіном, або лише спонгіном. Належать губки кліони (родина Сііопійае), які мають здатність просвердлювати ходи у вапняках, келих Нептуна (Роіегіоп Неріипі), рогові губки (родина ЗропдіШае), бодяги (родина 8роп§ШШае) та ін. У середній та нижній течії Дніпра помічено 7 видів бодяг: 8роп£і11а Іасизігіз, 8. їта^іііз та ін.
Клас Археоціати - переважно вимерлі морські тварини, що відрізняються від справжніх губок монолітним скелетом і відсутністю спікул.
Роль та значення губок
Тварин, що живляться губками, дуже мало. Лише в шлунку деяких риб і тюленів знаходили рештки губок, проте в харчовому раціоні й цих тварин вони не відіграють помітної ролі.
Між губками та іншими організмами спостерігаються різні форми симбіозу. Так, багато ракоподібних, деякі багатощетин- кові черви використовують тіло губок як схованку і захисток від ворогів. У тканинах губок відкладають яйця кільчасті черви, рачки, риби. У парагастральній порожнині губок ховаються на ніч креветки та бокоплави (явище коменсалізму).
Між губками, особливо прісноводними, і водоростями встановлюються досить тісні взаємовигідні зв’язки. Так, зоохлоре- ли, потрапивши з водою в тіло губки, не перетравлюються відразу, а певний час живуть у клітинах губки. Губка використовує кисень і поживні речовини, що синтезують водорості. Останні знаходять тут захисток, мінеральні солі та вуглекислий газ, що виділяється губкою при диханні. Це явище протокооперацй. Зелений колір прісноводних губок зумовлений саме наявністю в їхніх тканинах зоохлорел.
Безперервно пропускаючи через своє тіло воду, губки є чудовими біофільтраторами. Підраховано, що губка об’ємом близько 10 кв. см проціджує за добу від 100 до 2000 л води. Особливо важлива їх роль у процесах самоочищення прісних водойм від гниючих органічних решток, бактерій і дрібних планктонних організмів.
Губки вилучають розчинені у воді речовини, у тому числі й ті, що містять кремній та кальцій, які йдуть на побудову їхнього скелету.
Окремі види губок є об’єктами промислу. Це перш за все туалетні губки (8роп§іа о^ісіпаИз, Ніррозроп§іа соттипіз), поширені в теплих морях, їх скелет складається лише із густої сітки рогових волокон. Туалетна губка з античних часів використовується людиною для миття тіла. На сьогодні туалетні губки використовуються і як полірувальний та шліфувальний матеріал в ювелірній практиці та для виготовлення фільтрів. Висушену та розтерту на порошок бодягу використовують при ревматизмі як розтирання, як гомеопатичні ліки - при невралгії тощо.
Свердлячі губки кліони, проробляючи ходи у вапняковому субстраті, руйнують скелясті узбережжя, коралові рифи (їх відмерлі частини), пошкоджуючи черепашки устриць, завдають шкоди устричним господарствам. Є ці губки і в Чорному морі, зокрема, такі види, як Сііопа иазііиіса, С. ІоЬаІа.
Запитання і завдання. 1. Виділіть найістотніші ознаки типу Губки. 2. Які клітини утворюють пінако- та хоанодерму губок? 3. Порівняйте губки типів аскон, сикон та лейкон. Чим вони відрізняються? 4. На які класи поділяють губок та які їхні найхарактерніші особливості? 5. Яку роль відіграють губки у водних екосистемах? Яке вони мають значення для людини? 6. Вивчіть видовий склад і розселення бодяг у навколишніх водоймах. 7. Проведіть спостереження за фільтраційною здатністю бодяги.
ТИП КИШКОВОПОРОЖНИННІ - СОЕІ.ЕМТЕКАТА
Кишковопорожнинні, як і губки, дуже давні тварини: рештки безскелетних медузоподібних організмів знайшли у відкладах вендського періоду протерозойської ери. Близькі до сучасних гідроїдні відомі з ордовика. На думку більшості вчених (Е. Гек- келя, 1.1. Мечникова та ін.), низько організовані багатоклітинні, до яких належать і кишковопорожнинні, походять від колоніальних джгутикових.
Як і губок, кишковопорожнинних у середні віки відносили до зоофітів. Проте вже у XVIII ст. спочатку французький лікар Пейсоннель (вивчав коралові поліпи), а потім і швейцарський натураліст Трамбле (уперше вивчив і описав гідру) довели тваринну природу кишковопорожнинних. Усередині XIX ст. зоолог Лейкарт виділив із типу 2оор1іу1;а як самостійний тип Кишковопорожнинні.
Тип Кишковопорожнинні об’єднує близько 9000 видів, з них у Чорному морі - 44 види. Населяють гідросферу, здебільшого моря й океани, проникаючи на глибину до 8-10 тис. м (окремі види медуз, актинії). Є й прісноводні форми. Більшість кишковопорожнинних веде прикріплений спосіб життя, деякі входять до складу зоопланктону.
Тіло цих тварин складається з двох шарів: зовнішнього - епідерми, який розвивається з ектодерми, і внутрішнього - гас- тродерми, що розвивається з ендодерми; між ними міститься майже позбавлений клітин шар - мезоглея. Наявна лише одна кишкова (гастральна) порожнина, яка відкривається назовні ротом. Симетрія тіла - радіальна, що пов’язано з водним, сидя- чоприкріпленим способом життя.
Більшості представників типу властиві дві життєві форми: поліп і медуза. У поліпів тіло циліндричної форми, на верхньому кінці розташований рот, оточений щупальцями. Поліпи часто утворюють колонії. Ведуть нерухомий або малорухомий спосіб життя. Медузи - поодинокі, вільно плаваючі, рухомі організми. їхнє тіло має форму дзвону із щупальцями по краях. У життєвому циклі кишковопорожнинних спостерігається чергування нестатевого (поліпи) і статевого (медузи) поколінь - метагенез або часткова редукція одного з цих поколінь - гіпоге- нез. Розміри широко варіюють від кількох міліметрів до кількох метрів. Так, медуза ціанея має зонтик до 2 м у діаметрі, а щупальця - до ЗО м завдовжки. Багатьом кишковопорожнинним властиве яскраве забарвлення. Це в поєднанні зі своєрідною формою надає їм особливої краси.
Тип Кишковопорожнинні ділиться на 3 класи: 1. Гідроїдні (Нусігогоа); 2. Сцифоїдні медузи (8 с у р Ь о г о а); 3. Коралові поліпи (А п І Ь о г о а).
КЛАС ГІДРОЇДНІ - НУБК020А
Гідроїдні - поодинокі та колоніальні форми, що зустрічаються переважно в морях, хоч є серед них і прісноводні. Характерне чергування нестатевого покоління - поліпа і статевого - медузи. Гідри (Нусіга оііеасіів) (рис. 31), які населяють прісні водойми, представлені лише поодинокими поліпами. Водяться прісноводні гідри в заводях річок, озерах і ставках, здебільшого на нижньому боці листків водяних рослин, на підводному камінні. Розміри тіла - до 5 см, щупалець - до 25 см. До субстрату прикріплюються нижньою частиною тіла - підошвою. На протилежному кінці розташований рот, який веде в кишкову, або гаст- ральну (гр. §азІгоз - «шлунок») порожнину. Рот оточений 6-12 щупальцями. Стінка тіла складається з епідерми та гастродер- ми; між ними розташована тонка базальна перетинка - безструктурна речовина (мезоглея не розвинена).
Клітини
епідерми гідри неоднорідні. Це здебільшого
епітеліально-м’язові
клітини,
зовнішня частина яких утворює покриви,
а внутрішня витягнута в скоротливе
м’язове волокно. В епідермі під епітелієм
розташовані нервові клітини (нейрони)
зірчастої
форми, з’єднані між собою відростками.
Це примітивної будови дифузна нервова
система (дифузний
плексус),
яка є основою для рефлекторної діяльності
тварин. Між епітеліально- м’язовими
клітинами, особливо на щупальцях і
навколо рота, розташовані жалкі
клітини
(кнідоцити),
які служать для захисту та нападу.
Кожна така клітина має капсулу, заповнену
отруйною речовиною. Тут же міститься
спірально закручена нитка з чутливим
відростком - кнідоцилем.
При подразненні кнідоциля капсула
скорочується, назовні викидається
жалка нитка, вкрита шипами, і ранить
тіло жертви, паралізуючи її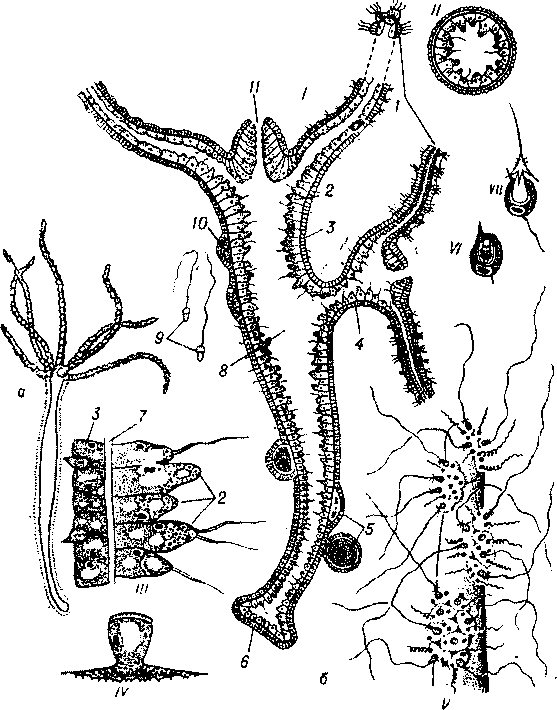
Рис. 31. Прісноводна гідра - загальний вигляд (а) і внутрішня будова (б):
І - поздовжній зріз; II - поперечний зріз; III - частина поперечного зрізу при сильному збільшенні: IV - епітеліально-м’язова клітина; V - кінець щупальця з викинутими жалкими клітинами; УІ-УІІ - жалкі клітини: І - щупальця: 2 - ентодерма; 3 - ектодерма; 4 - молода гідра, що відпупковується; 5 - яйце на різних стадіях розвитку; 6 - підошва; 7 - опорна пластинка: 8 — гастральна порожнина; 9 — сперма- тозоони; 10 - сім’яники; 11 - ротовий отвір.отрутою. Такі великі капсули, що служать для пробивання покривів тіла й ураження здобичі, називаються пенетрантами. Інші ж, менші, - вольвенти — мають короткі спірально закручені нитки, які обвиваються навколо різних виступів на тілі здобичі й утримують її. І, нарешті, видовжені капсули - глюти- нанти - мають довгі липкі нитки, що служать для утримання жертви. Біля основи епітеліально-м’язових клітин є також дрібні недиференційовані інтерстиціальні клітини (і-клітини), які можуть перетворюватися на інші типи клітин епі- та гастродер- ми, замінюючи загиблі.
Гастродерма складається з епітеліально-м’язових і залозистих клітин; останні виділяють травний сік із ферментами в кишкову порожнину, де частково відбувається травлення. Епітеліальні частини епітеліально-м’язових клітин мають 2-5 джгутиків і здатні утворювати псевдоніжки, якими захоплюють дрібні частки їжі, що перетравлюється всередині клітини. Джгутики сприяють переміщенню вмісту гастральної порожнини. Отже, у цих тварин поєднуються внутрішньоклітинне та порожнинне травлення. Живляться вони переважно дрібними планктонними найпростішими, рачками і коловертками. Здобиччю гідр стають і більш високоорганізовані тварини - черви, мальки риб. М’язові відростки епітеліально-м’язових клітин гіподерми діють як антагоністи м’язових відростків епідерми. Разом вони забезпечують рух щупалець, скорочення тіла; за їх допомогою гідра може повільно пересуватися на підошві або «крокувати» за допомогою щупалець і підошви. Дихають гідри всією поверхнею тіла. Продукти обміну речовин осмотично видаляються в гастральну порожнину, з якої разом із неперетравленими рештками їжі через ротовий отвір викидаються назовні.
Розмножуються гідри як безстатевим, так і статевим способом. За сприятливих умов на тілі гідри утворюються 1-3 бруньки, які з часом відокремлюються від материнського організму. Це - безстатеве розмноження. Восени гідри розмножуються статевим способом. Під епідермою утворюються гонади (гр. §опе - «народження»), в яких дозрівають яйцеклітини та сперматозоїди. Після запліднення яйцеклітини гідра гине. Навесні із заплідненого яйця розвивається новий поліп. Отже, гідра - поліп, що втратив медузоїдне покоління; не має вона і вільноплаваючої личинки.
У гідри чудово виражена здатність до регенерації - відновлення втрачених частин. Навіть 1/200 частина тіла може дати початок новому організмові. Перші досліди по вивченню регенерації у гідри були проведені понад 200 років тому Трамбле. Термін «регенерація» запропонував у 1712 р. Реомюр. Основні закономірності регенерації було сформульовано понад 100 років тому Ч. Дарвіном у вигляді двох правил. По-перше, тварини, що перебувають на нижчому ступені розвитку, значно легше відновлюють утрачені частини, ніж високоорганізовані. По-друге, здатність до регенерації значно вища в молодому віці (або на ранніх стадіях розвитку), ніж у зрілому. На даному етапі розвитку науки встановлено, що в основі регенерації лежать закономірності, подібні до таких при нормальному розвитку. У зв’язку з цим про регенерацію можна говорити як про явище повторного розвитку. Найбільша подібність регенерації з явищами нормального розвитку виявляється в тих випадках, коли регенерація відбувається за рахунок малодиференційованих клітин, що й характерно для гідри. Спеціальні дослідження із застосуванням методів мічених атомів та електронної мікроскопії засвідчили, що при регенерації відбувається дедиференціювання клітин, за якого клітини втрачають свої специфічні ознаки, спрощується їх будова, вони набувають ознак, властивих клітинам ембріонального типу. Де- диференційовані клітини здатні до активного розмноження.
У водоймах найчастіше зустрічаються такі види: стебельчаста (Нуйга оііцасііз), звичайна (Н. Vи^§а^І8) та інші. У зеленої гідри (Скіогокуйга иігШіззіта) в ектодермі живуть одноклітинні водорості - зоохлорели. Вони перебувають у симбіозі з гідрою. Зоохлорели використовують вуглекислий газ і аміак, які виділяються клітинами гідри. Тому зелена гідра переносить забруднення води легше, ніж інші види гідр.
Серед гідроїдних є морські поліпи, які утворюють колонії з великою кількістю особин. До них належить обелія (ОЬеІіа §епісиІаіа), поширена і в Чорному морі, та кордилофора каспійська (СогйуІорНога сазріа), що зустрічається на опріснених ділянках Каспійського, Азовського, Чорного та інших морів. Колонія має вигляд невеликого деревця чи кущика. На спільному стовбурі колонії сидять окремі особини, або гідранти, які за будовою нагадують гідру. Гастральні порожнини гідрантів сполучаються між собою; їжа, захоплена одним із них, розподіляється по всій колонії. Епідерма поліпа виділяє органічну оболонку - теку, або перидерм, що надає йому більшої міцності. Поліпи розмножуються нестатевим шляхом - брунькуванням, проте дочірні особини не відокремлюються від материнського організму, утворюючи колонію з великої кількості особин.
На гілочках гідрополіпів утворюються видозмінені особини, позбавлені рота і щупалець, - бластостилі, з яких розвиваютьс
я
маленькі медузки. Вони відокремлюються від поліпів і активно плавають у воді. За своєю будовою гідромедузи подібні до сцифо- медуз, але невеликих розмірів (1-2 мм, рідше - кілька сантиметрів у діаметрі). Крім того, у гідромедуз є парус - тоненька кільцеподібна подвійна складка епідерми, розміщена по вільному краю дзвона, що звужує діаметр вустя дзвона і відіграє важливу роль у плаванні. При одночасному скороченні дзвона і паруса вода виштовхується з-під дзвона і медуза рухається опуклим боком уперед. Медуза росте, дозрівають її статеві органи - гонади, у яких з клітин ектодерми утворюються статеві клітини - яйцеклітини та сперматозоїди. Медузи - різностатеві. Розвиток заплідненого яйця відбувається поза материнським організмом; з нього спочатку формується паренхімула, що нагадує личинку губок, а потім планула. Вона спочатку плаває, потім прикріплюється до дна і перетворюється в маленького поліпа, який брунькується, росте. Отже, у гідроїдних поліпів життєвий цикл складається з двох поколінь, які чергуються: нестатевого - поліпа і статевого - медузи (явище метагенезу). Медузи виконують функцію розселення виду.
Прісноводні гідри існують лише у формі поліпа, ніколи не утворюючи медуз. Гідри та гідроїдні поліпи об’єднуються в підклас Гідроподібні (Нуйгоісіеа).
Д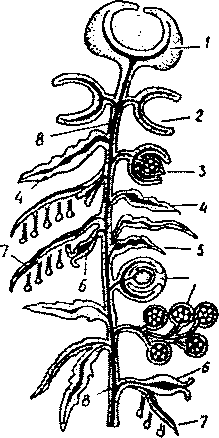 о
підкласу Сифонофори (8ір1іопор1іога)
належать
вільноплаваючі колоніальні морські
організми, що зустрічаються в морях і
океанах з високою солоністю води.
Відомо близько 170 видів. Колонія
складається з головного стовбура, на
якому розміщені особини (зооїди),
що мають різну будову і функції. Такі
колонії називаються поліморфними
(рис. 32).
о
підкласу Сифонофори (8ір1іопор1іога)
належать
вільноплаваючі колоніальні морські
організми, що зустрічаються в морях і
океанах з високою солоністю води.
Відомо близько 170 видів. Колонія
складається з головного стовбура, на
якому розміщені особини (зооїди),
що мають різну будову і функції. Такі
колонії називаються поліморфними
(рис. 32).
Як стовбур, так і зооїди складаються з епі-, мезо- та гастродерми зі спільною гастральною порожниною. Верхній сліпо замкнений кінець стовбура закінчується повітряним пухирем, що має назву пневматофор, залозисті клітини
Рис. 32. Схема будови сифонофори:
І - пневматофор; 2 - плавальний дзвін (некто- фор); З - статева особина (гонофор); 4 - покривна пластинка; 5 - пальпон; 6 - годуючий поліп (гастрозоїд); 7 - арканчпк; 8 - стовбур колонії.
якого виділяють газ, близький за своїм складом до повітря. Пневматофор буває дуже маленький - 1-20 мм, але іноді досягає 20-30 см. Це своєрідний гідростатичний апарат. Якщо він заповнений повітрям, то сифонофора плаває біля поверхні моря, коли ж повітря виходить, то колонія занурюється на глибину. Пневматофор може регулювати кількість повітря, випускаючи його крізь пору і знову утворюючи за допомогою залозистих клітин. Під міхуром на стовбурі розміщені плавальні дзвони, або нектофори - недорозвинені медузоїдні особини, що не мають щупалець і рота. їх буває від 10 до 400. Постійно скорочуючи свої парасольки, вони разом із пневматофором забезпечують рух колонії.
Важливу роль у сифонофора відіграють гастрозоїди, кожен з яких має довге щупальце - арканчик із численними жалкими клітинами. Арканчик захоплює і паралізує здобич, яка потім направляється до широкого рота гастрозоїда. Отже, гастрозоїди виконують функцію живлення.
На стовбурі сидять також пальпони і цистозоіди - це ніби спрощені гастрозоїди з нерозгалуженим щупальцем. Допускають, що вони виконують функцію виділення і чуття. Крім того, колонія обов’язково має статеві медузоїди, або гонофори, у яких розвиваються статеві залози. Причому медузоїди бувають і чоловічі, і жіночі.
Із заплідненого яйця розвивається личинка планула, а з неї
дорослий організм, окремі зооїди на якому утворюються, шляхом брунькування.
Преставником цього підкласу є фізалія, або «португальський кораблик» (Ркузаііа агеікиза), довжина стовбура в якої становить більше одного метра, а арканчиків - близько 10 м. Фізалія належить до плейстонних тварин, що рухаються під впливом не лише водного, а й повітряного середовища. Сифонофор у них великий (близько ЗО см), завжди перебуває на поверхні води і служить парусом. За допомогою вітра великі фізалії можуть рухатися зі швидкістю до 26 м/сек.
КЛАС СЦИФОЇДНІ МЕДУЗИ - 8СУРН020А
До цього класу належить близько 200 видів, поширених виключно в солоних водоймах. Мають складнішу будову, ніж поліпи, оскільки це плаваючі, рухливі організми. Тіло їх прозоре, у формі дзвона, зі щупальцями по краях, які мають жалкі клітини. Медузи на 98% складаються з води (рис. 33). Зовнішній опук-
а
а 6
Рис. 33. Схема будови сцифомедузи Аигеїіа аигіїа - вигляд з орального боку (а), розріз через середину тіла (б):
1 - ротові лопаті; 2 - ротовий отвір; 3 - щупальця; 4 - ропалій; 5 - кільцевий канал; 6 - радіальний канал; 7 - гонада; 8 - гаотральні нитки; 9 - шлунок; 10 - ексумбрела; 11 - субумбргла; 12 - мезоглея.
лий бік тіла медузи називається ексумбрелсю, внутрішній увігнутий - субумбрелою.
Нервова система більш складна порівняно з гідромедузами. Крім дифузного плексуса, у них є 2 нервових кільця по краю дзвона - зовнішнє і внутрішнє, а також скупчення нервових клітин - гангліїв (гр. §ап§Ііоп - «вузол»), розташованих біля органів чуття. Органи чуття знаходяться по краю дзвона в ропаліях - видозмінених щупальцях. Кожен із ропалій несе один стато- цист (гр. зіаіоз - «стоячий», кизііз - «скриня») - орган рівноваги - та кілька очок. За допомогою статоцистів медузи також сприймають інфразвуки, що виникають від тертя морських хвиль об повітря. Цим пояснюється здатність медуз заздалегідь відчувати початок шторму і завчасно відпливати від берегів. На основі цієї властивості медуз учені-біоніки сконструювали прилад, який дає змогу передбачати наближення шторму за 15 годин.
З нижнього боку дзвона в медуз розташований рот, оточений ротовими лопатями, рясно вкритими жалкими клітинами. За допомогою лопатей медузи захоплюють здобич. Рот веде в шлунок, що утворює 4 кишенеподібних виступи, гастродерма яких містить переважно залозисті й травні клітини (гастраль- ні нитки). Тут відбуваються процеси порожнинного та внутрішньоклітинного травлення. Від шлунка до країв дзвона розходиться система радіальних каналів, з’єднаних із кільцевим, розташованим по краю дзвона. Стінки каналів вистелені епітеліальними клітинами з джгутиками, які забезпечують рух продуктів травлення, отже, виконують в основному транспортну функцію. Шлунок разом із каналами утворює характерну для медуз гастррваскулярну систему, що виконує функції травної і кровоносної систем. Живляться медузи планктоном, у тому числі й мальками риб.
Завдяки м’язовим волокнам краї дзвона скорочуються, вода виштовхується з-під нього, і медуза рухається в протилежному напрямку {реактивний рух).
Медузи роздільно статеві. Статеві залози - гонади - розвиваються із гастродерми шлунка. Дозрілі статеві клітини виходять назовні через рот. Із запліднених яєць розвиваються вільноплава- ючі личинки - планули, які з часом прикріплюються до субстрату і перетворюється на невеликі окремі поліпи - сцифістоми. Шляхом поперечного поділу (стробуляції) поліп утворює дископодібні личинки - ефіри, що вільно плавають; з них розвиваються медузи.
Отже, у сцифомедуз, як і в гідроїдних поліпів, спостерігається чергування поколінь (метагенез): статевого — медузи і нестатевого - поліпа. Проте, на відміну від гідроїдних, у сцифомедуз поліпоїдна стадія розвинена слабо: сцифістоми дуже дрібні, вони ніколи не утворюють колоній. Гідромедузи невеличкі, менше сантиметра в діаметрі, а сцифомедузи (медузоїдна стадія) досягають чималих розмірів (аурелія буває в діаметрі до 40 см). У Чорному морі виявлено 3 види медуз: аурелія, або вухата медуза (Аигеїіа аигііа), коренерот {КНігозіота риїто) і сидяча медуза {Ьисегпагіа сатрапиіаіа) (рис. 34).
В останні роки зафіксовано різке збільшення їх чисельності. Якщо в 1960-ті роки біомаса аурелії в Чорному морі, за підрахунками вчених, становила в середньому 700 тис. т, то через два десятиліття їх ртало вже більше 40 млн. т. Однією з причин спалаху чисельності медуз є скорочення запасів планктоноїд- них риб, які конкурують у живленні.
Коренерот отруйніший за аурелію; у людини він викликає опіки, які не проходять протягом кількох годин. Щоб уникнути опіку при зіткненні з цією твариною, її потрібно відштовхувати від себе за опуклу частину дзвона, де жалких клітин немає. У симбіозі з коренеротом перебувають мальки ставриди, які у випадку небезпеки знаходять притулок між ротовими лопатями та під дзвоном.Р
коренерот,
ис. 34. Медузи: а - аурелія; б - «компасна» медуза; в -До цього класу належать морські тварини виключно поліпоїдної форми, поширені здебільшого в теплих морях. Відомо понад 6000 видів, з них у Чорному морі - чотири, в Азовському - один.Коралові поліпи здебільшого утворюють великі колонії, для підтримування яких у процесі еволюції розвинувся міцний вапняковий (рідше роговий) внутрішній або зовнішній скелет. Існують м’які корали, наприклад, альціонп (Аісуопіитп). Є серед коралових і поодинокі форми, зокрема, актинії (Асііпіагіа), які не мають скелета. Висота окремих особин - від кількох міліметрів до 1,5 м. У життєвому циклі стадія медузи відсутня.
Форма тіла особин циліндрична. У колоніальних форм нижній кінець окремого поліпа прикріплюється до загального тіла ко лонії - ценосарку, а в поодиноких за допомогою підошви - до субстрату. На протилежному кінці тіла знаходиться ротовий діиск, оточений щупальцями з жалкими клітинами, кількість яких дорівнює 8 (у восьмипроменевих коралів) або є кратною 6 чи 4 (у зоантарій). У центрі ротового диска є рот, який веде в ектодермальну глотку, вкриту війчастим епітелієм. Вієчки постійно рухаються і женуть воду в кишкову, або гастральну, порожнину, розділену на камери перегородками (септами), кількість яких відповідає кількості щупалець. На перегородках є потовщення - мезентеріальні нитки, що несуть численні залозисті клітини, які виділяють травні ферменти.
У коралових поліпів добре розвинена мезоглея; під епідермою, яка складається зі справжніх епітеліальних клітин - шар м’язових клітин (відбулася диференціація м’язових та епітеліальних клітин). Нервова система в них також розвинена краще, ніж у гідроїдних, хоч і має вигляд дифузного плексуса.
Живляться коралові поліпи як фільтратори - планктоном, органічними рештками, що є у воді. Жертвами актиній (вони переважно хижаки) стають риби, рачки. Деякі поєднують обидва типи живлення. Всі коралові поліпи здатні поглинати розчинені у воді органічні речовини клітинами епідерми, а також живитися за рахунок симбіотичних водоростей.
Розмножуються нестатевим і статевим способами. Нестатеве розмноження найчастіше відбувається шляхом брунькування, рідше - поперечного та повздовжнього поділу окремих поліпів, а також лацерації - відокремлення від поліпа шматочка або групи клітин, з яких розвивається нова особина.
Під час статевого розмноження в гастродермі утворюються гонади; статеві продукти через рот викидаються назовні, де йвідбувається запліднення. Є серед них роздільностатеві та гермафродити. Із заплідненого яйця розвивається личинка плану- ла, яка з часом прикріплюється до субстрату і перетворюється на поліп. Чергування поколінь (отже, і стадія медузи) відсутнє.
До класу Коралові поліпи належать два сучасних підкласи: 1. Альціонарії, або Восьми- променеві корали (О с 1; о с о г а 1 1 і а); 2. Зоанта- рії (2 о а п іа г і а) (рис. 35).
Рис.
35. Коралові поліпи:
а
- частина колони черпоного корала; 1
-
розітнуті поліпи; 2
- канали; З
- вапняковий стержень; б
-
актинії, які сидять на черепашці,
зайнятій раком-самітником; 1
-
акоішії.
З о а н т а р і ї мають нерозгалужені щупальця, кількість яких (так само і септ) кратна 6. Вони або взагалі не мають скелета (актинії), або ж характеризуються масивним зовнішнім вапняковим скелетом (мадрепорові корали - Майгерогагіа), який будують із розчинених у воді солей кальцію, мезоглея тонка. У Чорному морі мадрепорові корали відсутні, актинії представлені двома видами: Асііпіа е^иіпа та Асііпоікое сіаоаіа. Це невеличкі актинії висотою 3-4 см, вони мають біля 200 щупалець. Перший вид поширений і в Азовському морі.
Актинії поширені майже в усіх морях, проте надають перевагу теплим. Заселяють як прибережну зону, так і значні глибини (до 11 км). Для поширення деяких евритермних видів вирішального значення набуває фактор солоності води. Наприклад, у Середземному морі є 41 вид актиній, у Чорному - 2, Азовському - 1, а в Балтійському їх немає зовсім, оскільки воно значно опріснене. Актинії відрізняються різноманітністю форм і кольорів. Важко знайти двох тваринок однакового забарвлення, яке часто різноколірне: темнобордовий тулуб і лілові щупальця; оливковий із червоними плямами тулуб і рожеві щупальця і т.д. Тіло актиній прозоре і видається відлитим із кольорового скла.
Цікавим є явище симбіозу актиній Айатзіа з раком-саміт- ником. Від такого співжиття мають вигоду обидві тварини: рак переміщує актинію, а вона його захищає від ворогів (протоко- операція).
У тропічних і субтропічних частинах Атлантичного, Індійського і Тихого океанів поширені рифоутворюючі коралові поліпи - мадрепорові. Для свого існування вони потребують значного вмісту солей (не нижче 3,5%), температуру води - понад 20,5°С. До того ж можуть жити лише в чистій прозорій воді, насиченій киснем. Тому найчастіше оселяються в зоні прибою, опускаючись на глибину до 50 м. Мадрепорові корали - колоніальні форми. Розмножуючись шляхом брунькування, вони розростаються й утворюють коралові рифи. У нижній частині колонія нежива, складається лише з мінеральних скелетних утворень, тоді як зверху продовжують наростати нові корали. Розрізняють берегові, бар'єрні коралові рифи та коралові острови (атоли). Найбільш переконливо причину утворення коралових рифів пояснив Ч. Дарвін (1842), вважаючи, що першою стадією є береговий риф, який внаслідок поступового опускання дна океану перетворюється на бар’єрний. Найбільшим на планеті є Великий Бар’єрний риф, що тягнеться на 1400 км уздовж східного узбережжя Австралії. Коли острів внаслідок тектонічних процесів повністю занурюється у воду, на його місці утворюється лагуна, а бар’єрний риф перетворюється на атол. Загальна площа коралових рифів становить близько 600 тис. кв. км. Коралові рифи утворюють характерні біоценози моря, головним компонентом яких завжди є певні види мадрепорових коралів. У заростях коралів знаходять притулок і їжу численні водорості, черви, молюски, ракоподібні, голкошкірі, риби. Коралові рифи характеризуються високою біологічною продуктивністю. Первинна продукція в екосистемі рифу становить від 50 до 300 г сирої біомаси на 1 кв. м за добу.
В останні десятиліття спостерігається нашестя на коралові рифи морської зірки тернового вінця (Асапіказіег ріапсі), яка, поїдаю- чи коралові поліпи, порушує структуру екосистеми. Однією з причин масового розмноження морської зірки є вилов туристами з метою колекціонування її природного ворога - варонії тритоніс, молюска з красивою черепашкою. Тут явне порушення рівноваги (у результаті діяльності людини) між взаємопов’язаними компонентами біоценозу в ланцюгах живлення: черевоногі мрлюски —» морські зірки —» коралові поліпи. Іншою причиною деякі вчені вважають забруднення морів, що призвело до загибелі дрібних хижаків, які раніше знищували велику кількість личинок тернового вінця. Кількість зірок на рифах, які омиваються чистою водою, невелика, але вона різко зростає в зонах забруднення.
Восьмипроменеві корали, або альціона- р і ї, мають 8 щупальців, 8 септ; скелет внутрішній, заглиблений у мезоглею, може бути як вапняковий, так і роговий. До підкласу належать горгонарії (Сощопасеа) з роговим скелетом, який містить значну кількість йоду (використовується в китайській медицині) і просякнутий вапном. Серед них відомий благородний, або червоний корал (Согаїїіит гиЬгит), твердий скелет якого забарвлений у рожевий чи цегельно-червоний колір. У Чорному морі є два види восьмипроменевих коралів: морське перо (Уігциіагіа тігаЬіІіз) та Раскусегіапікиз зоШагіиз.