Продукты жизнедеятельности протопласта
В процессе жизнедеятельности протопласта возникают разнообразные вещества, получившие обобщенное .название оргастических (от греч. «эргастикос» — деятельный). Они образуются непосредственно в цитоплазме и отчасти сохраняются в пей в растворенном виде либо в форме включений. В значительно больших количествах оргастические вещества концентрируются вне протопласта, образуя оболочку клетки. Друтя часть накапливается в клеточном соке вакуоли в виде растворов или откладывается в виде включений. Природа и основные функции эргастических вещееi в различны. Главнейшие из них: простые белки (протеины); некоторые ушеводы, в частности глюкоза, сахароза и крахмал или близкий к нему инулин, а также целлюлоза; запасные жиры и жироподобные вещества, продукты вторичпо1 о метаболизма — питпиды, полифенолъпые соединения, алкалоиды, изопреипроизводиые и др. К эркь стическим веществам ошосшся иноке обычный во многих растениях оксалат кальция. Вещества вторичного мемболиз- ма нередко химически связаны с сахарами и образуют так называемые гликози- лированные формы, или гликозиды. Гли- козиды отличаются от негликозилиро- ванных соединений прежде всего повышенной способностью проникать через биоло1 ические мембраны.
Почти все эр1 астичеекие вещества независимо от их природы в той или иной мере могут вновь вовлекаться в процессы активного метаболизма клетки. Поэтому деление этих веществ на ряд ipynn по их равнейшей функции в известной мере условно.
Важнейшая группа эр1 астических веществ — запасные ‘ вещества. Это протеины, перечисленные выше углеводы, исключая целлюлозу, и жиры. Их образование и формы накопления будут подробнее рассмотрены в разделе «Включения». Целлюлоза и близкие к пей по химической природе гемицеллюлозы и пектиновые вещества — структурные компоненты клеточной оболочки. Вторичные метаболиты выполняют различные функции, но главнейшая из них связана с «организацией» защиты от поедания растений животными, внедрения болезнетворных микрооршшзмов и т. д. Таким образом они являются группой защитных веществ. Оксалат кальция чаще всего рассматривается в качестве конечного продукта метаболизма, формы «захоронения» ненужных или вредных клетке веществ. Реутилизируетея, т. с. вторично используется в процессах активного метаболизма, оксалат кальция редко.
Большинство эргастических веществ
физиоло1 пчески акшвпо. Многие из них накапливаю 1ся в значительных количествах и имеют исключительное значение в хозяйственной деятельности человека и медицине. Общеизвестно многообразное использование целлюлозы, или клетчатки. В юхнике пищевой промышленно- сш и медицине широко используются крахмал, ппокоза и сахароза. Таппиды, или дубильные вещества, полифепольные соединения и пзопрепнроизводпые —основа для получения болыпнпава медицинских препараюв paeimejibiioio происхождения.
Наконец, oimcium еще один момент, связанный с '>pi аоическими веществами и важный для подютовкп будущих провизоров. Мпоше соединения откладываются в форме разного рода включений — крахмальных зерен, кристаллов и г. д., имеющих характерную форму. Это широко используется для распознавания, т. е. диагностики, объектов в процессе микроскопического анализа лекар- ci венного растительного сырья.
Цитоплазма
ЦиIонлазма -™ часть протопласта, заключенная между плазмалеммой и ядром. Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоилазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клсточшм о ме- щболизма. В цитоплазме растительных клеток имеются органоиды: небольшие тельца, выполняющие специальные функции,—пластиды, комплекс Гольджи, зп- доплазматическая сеть, митохондрии и т. д. В цитоплазме осуществляется большая часть процессов клеточного метаболизма, исключая синтез нуклеиновых кислот, происходящий в ядре. Цитоплазма пронизана мембранами —тончайшими (4—10 нм) пленками, построенными в основном из фосфолипидов и липопротеи- иов (рис. 7). Мембраны придают структуру всем органоидам клетки и ядру, отграничивают цитоплазму от клеточной оболочки и вакуоли и внутри цито-
Рис.
7. Модель биоло) ичеекой мембраны.
В
двойном лшшдпом слое (/) молекулы шшидон
паиранпепы своими шдрофильиыми концами
(обо- чпачеиы кружочками) к слоям бейкой
(2). В юнах шдрофиньиых пор (Л) шдрофильные
концы липидных молекул обращены дру!
к дру1
у. Белковые слон еоеюя! ич 01
дельных еубьедипиц. Пекоюрые субь-
единицы (4)
пересекаюI всю мембрану Оуппельиые
белки)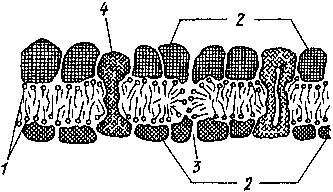
Яковлев
Г.П., Челомбитько В.А. БОТАНИКА
Одно из основных свойств мембран клетки — их избирательная проницаемость (полупротщаемость): одни вещества проходят через нее с трудом пли вообще пе проходят, дру1ие проникают легко. Избирательная проницаемость мембран создает возможность одновременного и независимою протекания различных биохимических реакций, нередко противоположных по направлению, в изолированных отсеках, образованных опдоплазматичеекой сетью.
Эпдоплазматическая сеть может быть гранулярной, т. е. несущей прикрепленные рибосомы, и гладкой (аграпулярнои) — лишенной рибосом. Гранулярная сеть вы-
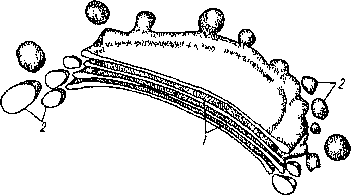
Рис 8. Схема строения комплекса Гольджи с отделившимися от них пузырьками Гольджи' 1 - цистерны, 2 - пузырьки
Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
Важнейшим свойством цитоплазмы, связанным прежде всего с физико-химическими особенностями гиалоплазмы, является ее способность к движению. В клетках с одной крупной вакуолью движение осуществляется обычно в одном направлении (циклоз) за счет особых органоидов — микрофиламентов, представляющих собой нити особого белка — актина. Движущаяся гиалоплазма увлекает пластиды и митохондрии. В молодых клетках с многочисленными мелкими вакуолями движение осуществляется струйчато. Интенсивность движения зависит от температуры, степени освещения, снабжения кислородом и т. д.
В цитоплазме, а также ядре, митохондриях и пластидах всегда находятся мельчайшие гранулы, выявляемые на ультратонких срезах под электронным микроскопом. Это рибосомы, представляющие собой частицы, образованные рибонуклеопротеидами. Располагаются рибосомы поодиночке, либо группами (полисомы), где отдельные составляющие связаны между собой нитевидной молекулой РНК. Рибосомы эукариотических организмов состоят из двух неравных по величине субъединиц, по-видимому, объединенных ионами Mg2+. Рибосомы нередко прикреплены к мембранам эндо- плазматической сети (гранулярная сеть) или свободно располагаются в гиалоплазме. Рибосомы (точнее, полисомы) — центры синтеза белка. '
Комплекс Гольджи (аппарат Гольджи) назван в честь итальянского ученого К. Гольджи, описавшего его. Комплекс состоит из отдельных диктгюсом (от греч. «диктион» — сеть, «сома» — i ело) и пузырьков Гольджи (рис 8). Диктио- сомы представляют собой стопку плоских, не соприкасающихся друг с другом дисковидных мешочков — цистерн, ограниченных мембраной. Часто диктиосомы переходят по краям в систему тонких ветвящихся трубок. Число диктиосом в растительной клетке обычно колеблется от одной до нескольких десятков. Пузырьки Гольджи отчленяются от краев дик- тиосомных пластинок или концов трубок и направляются обычно в сторону плаз- малеммы или тонопласта. Считается, что в диктиосомах синтезируются полисахариды. Пузырьки Гольджи транспортируют образовавшиеся полисахариды, которые у растений нередко участвуют в формировании клеточной стенки. Кроме того, комплекс Гольджи, по-видимому, участвует в формировании вакуолей.
Лизосомы (от греч. «лизис» — растворение, распад и «сома» - тело) ~ органоиды грибов и животных, отсутствующие в растениях. Представляют собой пузырьки, окруженные мембраной. Образуются в комплексе Гольджи. Содержат набор гидролитических ферментов. С их помощью осуществляется внутриклеточное пищеварение.
Микротрубочки представляют собой полые длинные цитоплазматические частицы диаметром 18-30 нм, видимые в электронный микроскоп. Предполагают, что благодаря активному перемещению микротрубочек осуществляются направленные движения пузырьков Гольджи. Нити веретена, появляющегося при делении ядра, также построены из большого числа микротрубочек.
Микрофиламенты (от греч. «микро» — малый и лат, «филаментум» — нить) — органоиды эукариотических, в том числе растительных клеток, образующих под цитоплазматической мембраной сплошные сплетения нитей белка-актина. Микрофиламенты способны к сокращению и непосредственно участвуют в изменении формы клеток, не обладающих твердой оболочкой. В растительных клет
ках микрофиламеш ы ответственны за цикл оз.
Митохондрии — neoi ьемлемая часть всех живых )укариошческих клеюк. Форма, вепичипа и их число посюяшю ме- НЯЮ1СЯ. Число митохондрий варьирует oi нескольких десятков до coien, особенно мною их в секреторных тканях растений. Размеры этих ортанондов не превышают 1 мкм, но форме они чаще всею эллин 1ические пли окрутлые. Снаружи миюхондрин окружены оболочкой, состоящей из двух мембран, которые не связаны с эпдоплашатической сегыо цитоплазмы. Внутренняя мембрана образует выросты в полость митохондрии в виде пластин или трубок, называемых кристами. Кристы бывают различных типов. Пространство между кристами заполнено однородным прозрачным веществом — матриксом митохондрий (рис. 9). В матриксе встречаются рибосомы, подобные по величине и строению рртбосо- мам прокариотических клеток, и собственная митохондриальная ДНК, заметная под электронным микроскопом в виде тонких нитей.
Основная функция митохондрий состоит в обеспечении энергетических потребностей клетки. Здесь синтезируются богатые энергией АТФ и АДФ путем реакций окислительного фосфорилирова- ния. Энергия, запасаемая в молекулах АТФ, получается в результате окисления в митохондриях различных энергетически богатых веществ, главным образом сахаров.
Митохондрии способны к независимому от ядра синтезу своих белков па собственных рибосомах под контролем митохондриальной ДИК. Митохондрии образуются только от митохондрий путем деления. Согласно теории эндосимбиоза, митохондрии наряду с пластидами являются прокариотическими эпдосим- биоптами эукариотической клетки.
Пероксисомы. Эти оргаиеллы обнаружены с помощью электронного микроскопа в большинстве типов клеток. Они представляют собой чаще всего сферические или эллипсоидальные образования диаметром 0,2—1,5 мкм. Функции перок- сисом зависят от типа клетки. Подобно пластидам и митохондриям, перокси-
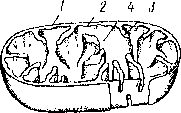
Яковлев
Г.П., Челомбитько В.А. БОТАНИКА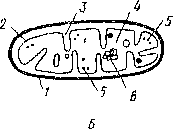
I — наружная мембрана оболочки мшохешдрий, 2 - внутренняя мембрана, 3 - криста, 4 - матрикс мик>- хондрпи, 5 - миюхонцриальныи рибосомы (оишча- Ю1ся oi цитилачма шческих рибосом), 6 - пип. ми I охопдриа ui.Hofi ДНК
сомы до пзвеспюй степени авюпомпы.
II иимиды. Плазмиды представляют собой кольцевые двуцепочечпые молекулы ДНК, существующие в большинстве изученных клеток в автономном, не связанном с хромосомами состоянии. Они являются внехромосомными факторами наследственности и интенсивно используются в генетических исследованиях, особенно в генетической инженерии, в качестве молекулярных переносчиков чужеродной ДНК. Наиболее изучены бактериальные плазмиды.
Пластиды характерны только для растений. Они не найдены у грибов и у большинства животных, исключая некоторых фотосинтезирующих простейших. В зависимости от окраски, связанной с наличием тех или иных пигментов или их отсутствием, различают три основных типа пластид: хлоропласты (зеленого цвета), хромопласты (желтою, оранжевого или красного цветов) и лейкопласты (бесцветные). Обычно в клетке встречаются пластиды только одною типа. Однако установлено, что одни типы пластид могут переходить в другие.
Пластиды — относительно крупные образования клетки. Самые большие из них — хлоропласты - достигают у высших растений 4—10 мкм длины и хороню различимы в световой микроскоп. Форма окрашенных пластид чаще всего липзо- видная или эллиптическая. В клетках встречается, как правило, несколько десятков пластид, но у водорослей, где пластиды нередко крупны и разнообразны по форме, число их иногда невелико (1 —5). Такие пластиды называются хрома- тофорамч. Лейкопласты и хромопласты могут иметь различную форму.
Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
Рис
10 Cj
роение
хлоропласта А
—
схема строения хлоропласта в обьемиом
изображении (трехмерная), Б
- схема среза через хлоропласт 1
—
наружная мембрана оболочки хлоропласта,
2 — внутренняя мембрана, З-ирома, 4
грана,
5 - тилдкойд граны, 6 - тиллкоид с, ром и,
7 - шпь иласшднои ДНК, 8
- рибосомы хлоропласта (отличаются от
цитоплазматических рибосом),
0--крахмальные зерна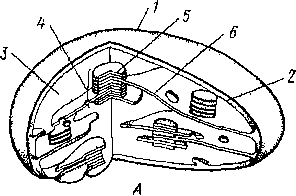
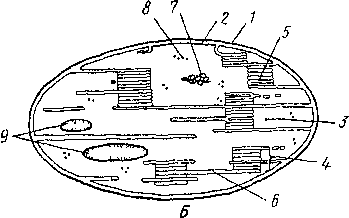
В строме хлоропластов содержатся ферменты и рибосомы, отличающиеся от рибосом цитоплазмы меньшими размерами Часто имеются один или несколько небольших зерен первичного ассимиляционного крахмала Генетический аппарат хлоропластов автономен, они содержат свою собственную ДНК Хлоро- пласты развиваются из проп шетид, которые способны к делению, независимому от деления клетки Полагают, что хлоропласты — это древние эндосимбионты — прокариоты, некогда поселившиеся в эукариотической клетке и ставшие ее часгыо.
Основная функция хлоропластов — фотосинтез. Центральная роль в этом процессе принадлежит хлорофиллу, точнее, его нескольким модификациям Световые реакции фотосинтеза осуществляются преимущественно в гранах, гем- новые — в строме хлоропласт. Как хлоропласгы, так и митохондрии способны синтезировать собственные белковые молекулы, так как содержат автономный генетический материал.
Помимо фотосинтеза, в хлоропласт ах осуществляется синтез АТФ и АДФ (фос- форилирование), синтез и гидролиз липидов, ассимиляционного крахмала и белков, откладывающихся в строме
Во многих случаях лейкопласты определяют синтез и накопление запасных питательных веществ, в первую очередь крахмала, иногда белков, редко жиров. Очень часто на основе лейкопластов возникают зерна вторичного запасного крахмала.
Яковлев
Г.П., Челомбитько В.А. БОТАНИКА
в них карошиоидов. Счишеюя, чю хро- моиласш — конечный 'лап и развитии пластид, иначе юворя, 'по сшрсющие хлоронласты и лейкопласты. Косвенное биоло/ическое ■значение хромопластов связано с яркой, привлекательной для животных окраской miioi их цветков и плодов.
Пигменты пластид
Пшмеш ы, локализующиеся и пластидах и участующие в процессах фотосии- хеза, прппадлежаг к трем хороню известным классам. Это х.юрофи.иы, каро- пшпоиды и фикоби.пшы. Все они входят в состав пигментных систем в виде хромопротеидов, т. с. иигмепт-белковых комплексов.
В основе строения хлорофиллов лежит так называемый Mg-иорфириновый скелет. Кроме того, имеются различные заместители, например спирт фитол, придающие молекуле хлорофилла способность встраиваться в липидный слой биоло1 ических мембран.
и
бакто-риохлорофиллов R
r~ R 4~~
различные радикалы
0
1
Ra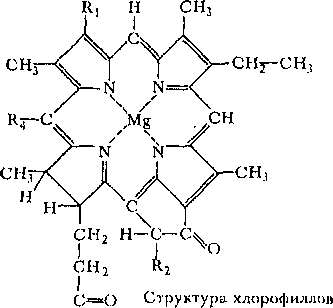
Хлорофиллы jieiKo извлекаются из растений водой и MOiyT использоваться в качес1ве естественных зеленых красителей в пищевой промышленности и медицине.
Каротиноиды — желтые, оранжевые или красные пигменты, синтезируемые растениями (а гакже бактериями и грибами). Каротиноиды отчасти выполняют роль дополнительных фотосинте1 ических пигментов, но при этом мшут осуществлять и другие функции, с фотоеишезом не связанные. К каротииоидам о г нося гея широко распространенные каротины и ксантофиллы. По химической природе ото изопрепоидпые углеводороды, содержащие 40 углеродных атомов. Ксантофиллы — окисленные каротины. Особенно богаты каротинами зеленые листья некоторых растений (например, шпината), корнеплоды моркови, плоды шиповника, смородины, томата и др.
Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
н
н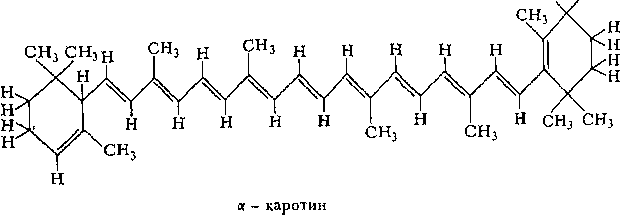
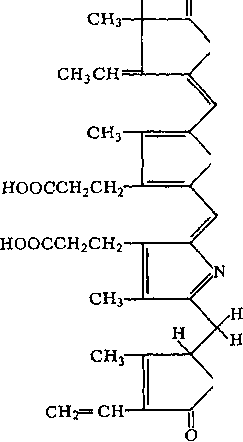
Фикобилины — пигменты багрянок, а также цианобактерий. По химической природе они представляют собой би- лины — вещества, близкие по составу к желчным кислотам, но соединенные с белками. Существуют красные фикобилины — фикоэритрины и синие — фико- цианины
.
N-H
фикоэритрин
N-H
;n-h
N—H
фикоциан
N-H
Н
О
N-H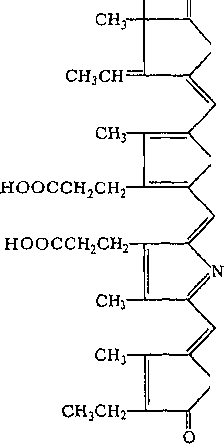
Фикобилины участвуют в фотосинтезе, доставляя поглощенную энергию света к молекулам хлорофилла.
Ядро
Ядро — обязательная и существеннейшая часть живой клетки всех эукариотических организмов. Это место хранения и воспроизводства наследственной информации, определяющей признаки данной клетки и всего организма в целом. Ядро служит также центром управления обменом веществ и почти всех процессов, происходящих в клетке. Из органоидов лишь митохондрии и пластиды в некоторой степени автономны и в части своих функций независимы от ядра. Клетки с удаленным ядром, как правило, быстро погибают. Отсутствует ядро в норме лишь в члениках ситовидных трубок флоэм
ыЯковлев
Г.П., Челомбитько В.А. БОТАНИКА
Чаще леею в клетках >укарио1 имеется лишь одно ядро, редко — два или не- екош>7со. Нормальное длительное существование в одной к не псе двух неслив- шихея ядер (дикаршт) характерно для мне» их 1 рпбов
Размеры ядра раишчпы, oi 2-3 до 500 мкм (у половых клеюк). Однако бе* специальной окраски ядро малозаметно. Форма ею чаще всею шаровидная или эллипсоидальная. В молодых кшмках оно ншимаст цетралыюе положение, по позднее обычно смещаеюя к оболочке, oi- юспяемое paciymeii вакуолью.
Общий план строения ядра одинаков у всех )укарио шческих орпшизмов. Снаружи оно окружено двойной мембраной — ядерной оболочкой, нрошпанной порами, па краях которых наружная мембрана переходит во внутреннюю. Содержимое ишерфазпото педелящеюся ядра составляет кариоп иана (или ядериый сок), близкая по структуре к шалоплазме. В кариоплазму погружены оформленные элементы: хроматин и чдрьиики, а гакже рибосомы. По химическому составу ядро отличается высоким содержанием ДНК. Большая часть ДНК клетки находится в ядре, в комплексах с ядерными белками. Основная масса ДНК сосредоточена в хромаI ине — особых нуклеопротеидиых нитях, рассеянных но всему ядру, В ядре заметны одно или несколько ядрышек. Подобно хроматину, ядрышки не имеют мембраны и свободно лежат в кариоплазме. Основная функция ядрышек — синтез некоторых форм РНК (в основном рибо- сомиой) и формирование предшественников рибосом.
Хромосомы -орпшоеды делящеюся клеточного ядра, являющиеся носителями тепов. Основу хромосом составляет одна непрерывная двухцепочечная молекула ДНК, связанная преимущественно с особыми белками (гистопамп) в пуклео- протеид. Строение молекулы ДНК обеспечивает запись наследственной информации. Управление синтезом белков осуществляется через посредство информационной РНК, образующейся в ядре под контролем ДНК и переходящей в цитоплазму (рис. 11). Хромосомы становятся видимыми во время клеточною деления и незаметны в интерфазе. Они образо-
Рис.
11 Схема, демонстрирующая роль различных
типов РНК в ядре и цитоплазме (по
Винчестеру)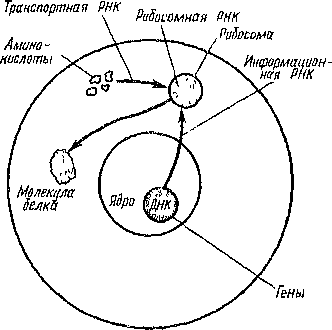
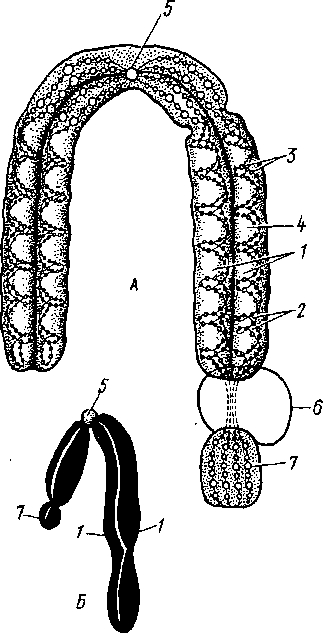
Ваны двумя сложенными но длине одинаковыми хрома гидами. Близ середины хромосомы имеют перетяжку, скрепляющую хроматиды,—цстпромеру (рис. 12). В клетках тела растений каждая пара хромосом представлена двумя гак называемыми I омоло1 ичпымп хромосомами, происходящими одна oi материнскою, а друтая oi отцовскою opiаппзма (двойной, или диплоидный, набор хромосом). Половые клетки содержат только одну из двух томолотичиых хромосом (та- плопдиый набор). Число хромосом у разных организмов варьирует oi двух до нескольких со теп. Все хромосомы в совокупности составляют хромосомный набор. Каждый вид имеет характерный и постоянный набор хромосом, закрепленный в процессе эволюции данного вида. Совокупность признаков хромосом- нот о набора (число, размер, форма хромосом), характерных для тою или иною вида, получила название кариотипа, Изменение хромосомною набора происходит юлысо в результате хромосомных и теиомных мутаций. Наследственное крагпое увеличение числа наборов хромосом получило название полиплоидии; не кратное гаплоидному изменению — апеуп- лоидии. Исследование кариотипа играет существенную роль при изучении систематики орт апизмов (кариосистематика).
Растения полип шиды часю характеризуются крупными размерами, повышенным содержанием ряда веществ,
1 — две хроматиды, т. е. половины хромосом, на которые она расщепляется продольно во время деления, 2 - хромонемы внутри хроматид, 3 — хромомеры, 4 — белковый матрикс хромосомы, 5 — первичная перетяжка с центромерой, 6 — ядрышко, 7 — спутник хромосомы
Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
Рис.
12. Строение хромосом (схема). А
— внутренняя структура; Б
- общий вид: