

)Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
0,1
мм
Рис.
24. Камбий и производные ткани. А
— схема вычлененной инициальной
клетки камбия, дающей начало проводящим
и волокнистым элементам флоэмы и
ксилемы; Б
— схема вычлененной инициальной клетки
камбия, дающей начало паренхимным
элементам сердцевинных лучей; из
аналогичных клеток возникает и паренхима
флоэмы и ксилемы; В
— поперечный срез через участок no6eiа,
захватывающий флоэму, камбий и ксилему;
Г
— продольно-радиальный срез через тог
же участок (см. пункт В);
Д -
продольно-радиальный срез через
сердцевинный луч (светлая сторона
— фдоэмпая часть, более 1емпая
— кси-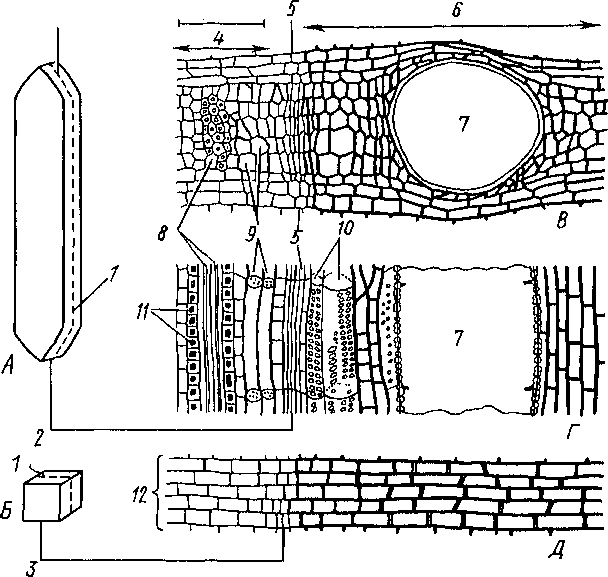
Лемпая):
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающих место поражения. Каллюсо- образовательиая способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразова- иие, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса—необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричпы по размерам и многогранны но форме. Межклетников между ними нех, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли мною- численные, мелкие, но под световым микроскопом обычно не заметны. Эргасти* ческие вещества, как правило, огсут
-Яковлев
Г.П., Челомбитько В.А. БОТАНИКА
сшукл. Пляс гид и миючондрнй мало и опп мелки
Клетки боковых мериаем различны по величине и форме Они примерно со- огветс1вуюг клеткам тех посюянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, гак и прозенхимпые инициал» Из паренхимных шшциалей образуется паренхима проводящих тканей, а из прозеихимиых — проводящие элементы (рис. 24).
ШЖРОВНМЬ I КАИН
Покровные ткани располагаю 1ся па границе с внешней средой. Большинегво из них состоит из плотно сомкнутых живых, реже мертвых клеток Покровные ткани выполняют барьерную роль, защищая внутренние ткани от высыхания и повреждения. Одна из функций покровных тканей — регуляция i азообмена и транспирации. Некоторые покровные ткани способны к всасыванию и выделению, активно регулируя скорость и избирательность проникновения веществ через них. Покровные ткани — барьер для проникновения патогенных микроорганизмов. Эволюциоппо покровная ткань — очень древнее образование, возникшее в момент выхода растений из водной среды на сушу. Подобно прочим постоянным тканям, покровные ткани возникают в процессе онтогенеза из меристем.
Принято различать первичные покровные ткани, образующиеся в результате дифферепцировки клеток первичных меристем. Это эпидерма и jгшбле- ма. Вторичная покровная ткань — перидерма — образуется из вторичной меристемы феллогена.
Эпидерма
Листья и молодые зеленые побеги, как чехлом, покрыты однорядной первичной покровной тканью — эпидермой. Изредка эпидерма многослойна. Эпидерма возникает из самого наружного слоя клеток - конуса нарастания. Это сложная ткань, поскольку ее клетки различаются по форме и отчасти по функциям. В частности, от остальных клеток эпидермы резко отличаются клс1ки, образующие устьица, и клетки трихом. Наружная поверхность клегок шидермы часю покры- ia слоем кутикх :ы или, реже, восковым палеюм различной юлщины Кугикула может достигать значительной юлщины, особенно у растений засушливых месю- обшлний. Нередко ее поверхность покрыта различного рода складками или бородавчатыми выростами Исключая успшчные щели, клетки шидермы плотно сомкнуты, т. е. отсутствуют межклетники. Главная функция эпидермы — регуляция пгзообмеиа и транспирации, i.e. иснарения воды растением. Газообмен и транспирация осуществляются преимущественно через устьица, по частично и через кутикулу. Кроме того, через поры и тяжи пектиновых веществ в наружных стенках клеточных оболочек эпидермы проникают вода и неорганические питательные вещества, что особенно характерно для водных растений. Инотда эпидерма выполняет необычные для этой ткани функции — гакие, как фотосинтез (у части водных растений), запасаппе воды (у некоторых пустынных растений), секрецию веществ вюричного метаболизма (ряд эфиромасличных растений).
Характер клеток эпидермы различен, большинство из них, получившее название основных клеток эпидермы, таблитчатые, разнообразны в очертаниях (рис 25). Боковые стенки нередко извилисты, чго повышает плотность их сцепления друг с другом, реже прямые. Эпидермальные клетки осевых органов п листьев многих однодольных сплыю вытянуты вдоль оси органа,
В основных клетках эпидермы обнаруживается тонкий постенный слой протопласта с мелкими редкими лейкопластами и ядром.
Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
Рис.
25. Эпидерма различных расюпий (в плане).
А
— хлорофшум (Chlorophytum),
представитель однодольных; Б
— плющ обыкновенный (Iledera
hrtix);
B
— iepaub душистая
(Pelargonium
qraveolens),
Г
—
шелковица
белая [Moms
alba); Б—
Г
— предсишиюли двудольных:
1
—
замыкающая клегка, 2
—
ус1ьнчпая
щепь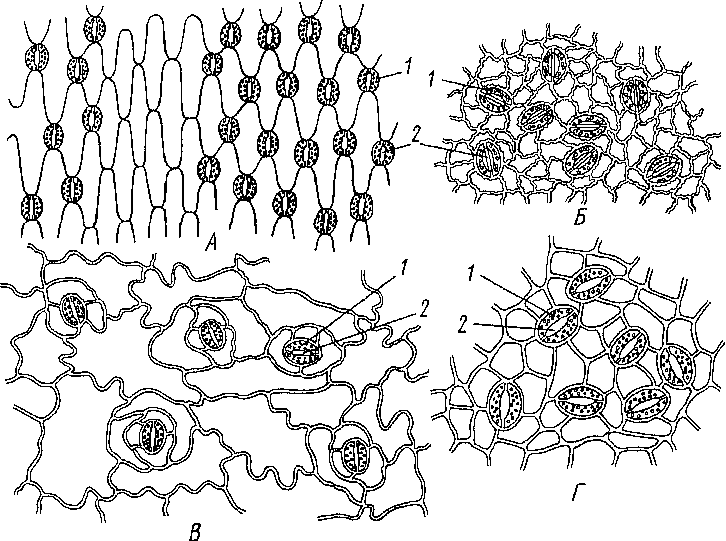
многих семян содержат полисахариды в виде слизи, которая при увлажнении набухает. Семена при этом легко приклеиваются к движущимся предметам и таким образом распространяются.
У некоторых растений под эпидермой расположена особая ткань — гиподерма. Она выполняв I от час ш механическую функцию, отчасти предохраняет растение от избыточною испарения.
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток и ус1ьичиой щели (своеобразного межклетника) между ними. Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен, Под щелыо располагается «дыхательная», или воздушная, полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим, получили название побочных или околоустьичных. Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устылчиыи аппарат (рпс. 26). Число побочных клсюк и их расположение относительно ус1ьичиой щели позволяют выделя1ь ряд уапьичпых питов (до 15). Их изучение получило название епю- матог рафии (от 1реч. «сюма» — усгьице). Данные сто мат oi рафии нередко используются в системашке растений для уюч» пения системашческою положения [аксонов, а также в фармако1 позии для мпкро- диапюстики лекарственною расiтельного сырья Наиболее час г о встречающиеся устьичиые типы показаны па рис. 27.
Для всех групп высших растений, исключая хвойные, весьма обычен аномо- цитиый тип усгьичною аппарата (oi греч. «апомос» — беспорядочный). Побочные клетки не отличаются от остальных клеток эпидермы. Диацитпый тип (от греч. «диа» — через) характеризуется только двумя побочными кле пса
-Яковлев
Г.П.,
Челомбитько
В.А.
БОТАНИКА
Рис.
26. Строение устьица (схема). А
— вид сверху; Б
- поперечный срез:
1
-
замыкающая клетка, 2
-
устьичная щель, 3
- побочные клетки, 4
- «дыхательная» полость, 5
— эпидермальные клетки, б
- кутикула, 7 - хлоропласты клеток
мезофилла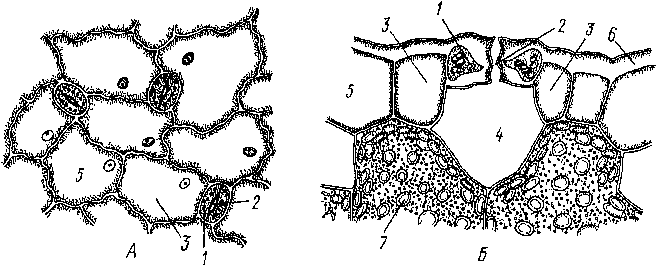
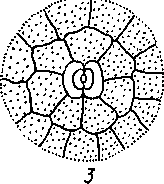
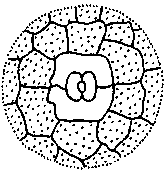
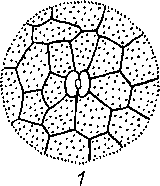
Рис.
27. Основные типы устьиц:
I
- аномоцитный (во всех группах высших
растений, исключая хвощи), 2-диа- цитный
(у лаггоропшков и цветковых), 3
- парацитный (у папоротников, хвощей,
цветковых и гпетопсид), 4
-
апшоцигный (только у цветковых), 5
- тетрацитный (главным образом у
однодольных), б - онцпклоцитный (у
папоротников, голосеменных и
цветковых)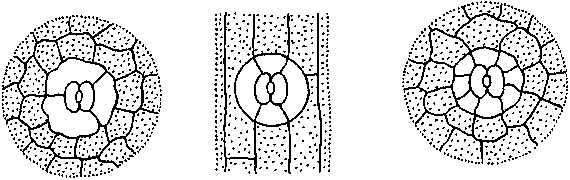
ми, общая стенка которых находится под прямым углом к замыкающим клеткам. Этот тип обнаружен у некоторых цветковых, в частности у большинства губоцветных и гвоздичных. При парацитном типе (от греч. «пара» — рядом) побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников, хвощей и ряда цветковых растений. Только у цветковых растений обнаружен анизоцитный тип (от греч. «анизос» - неравный). Здесь замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Однодольные характеризуются преимущественно тетра- цитным типом устьичного аппарата (от греч. «тетра» — четыре). При энциклоцит- ном типе (от греч. «киклос» — колесо) побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников, голосеменных и ряда цветковых. Наконец, для актиноцитного типа (от греч. «ак- тис» — луч) характерны несколько побочных клеток, радиально расходящихся от замыкающих клеток. Встречается этот тип только у цветковых. Расположение
Яковлев
Г.П., Челомбитько В.А. БОТАНИКА
Рис.
28. Кроющие, железистые волоски на
эпидерме и эмергсицы. А
- простые многоклеточные у картофеля;
Б
— простые одноклеточные у яблони; В
- ветвистые у коровяка; Г
— звездчатые у лоха (вид с поверхности);
Д
—
железистые с многоклеточной головкой
и ножкой у табака; Е
— последовательные стадии развития
{1—3)
жгучего омергенца у крапивы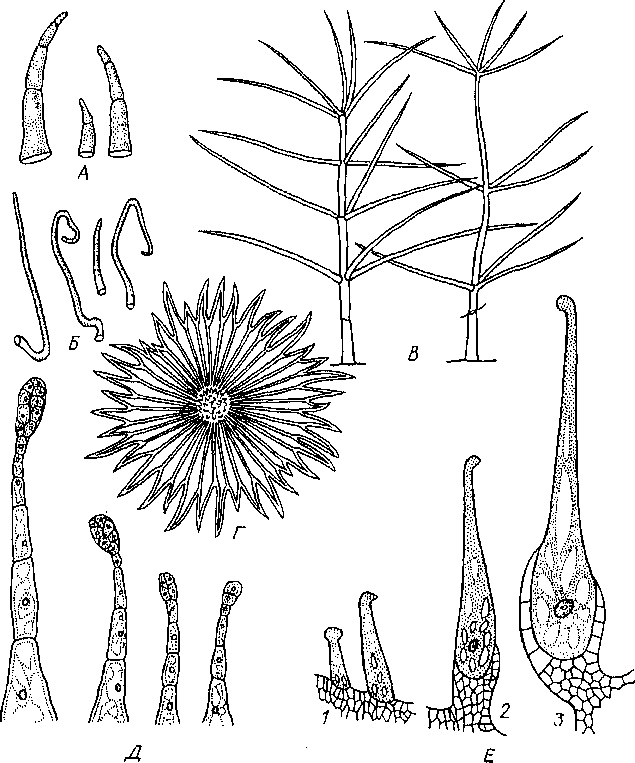
замыкающих клеток относительно прочих клеток эпидермы у разных видов неодинаково. В одних случаях замыкающие клетки находятся на одном уровне с эпидермальными, иногда выступают над ними или, напротив, залегают значительно глубже (погруженные устьица). Последнее наблюдается у растений, приспособленных к изменениям режима испарения (лист олеандра). Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками и называются устьичпыми криптами.
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растения и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 мм2 поверхности.
Яковлев
Г.П., Челомбитько В.А. БОТАНИКА
в изменении тургора принадлежит ионам калия Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов Первичный крахмал хлоропластов, превращаясь в сахар повышает концентрацию клеточного сока Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в состояние тургора Общая площадь устьичных отверстий составляет лишь 1 — 2% площади листа Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равною по площади открытой водной поверхности
Трихо мы (от греч «трихома» — волосы) у растений — это различные по форме, строению и функциям выросты клеток эпидермы — волоски, чешуйки, железки, нектарники Размеры трихом варьируют в значительных пределах Наиболее длинные трихомы (до 5 — 6 см) покрывают семена хлопчатника Трихомы могут быть живыми и отмершими и выполнять различные функции Они делятся на кроющие и же шзиапые Кроющие — это одноклеточные, многоклеточные, ветвисше и звездчатые волоски (рис 28) Железистые трихомы (железки и нектарники) являются элементами секреторных тканей Разнообразие кроющих трихом довольно велико Их строение и форму иногда используют для целей систематики В фармакогнозии особенности трихом эпидермы играют важную роль при микроскопическом анализе лекарственного растительного сырья Кроющие волоски образуюi на растении различной густоты опушение, предохраняющее от избыточной транспирации или изредка, напротив, ускоряющее ее На листьях они чаще возникают с гои стороны, где имеются устьица
Помимо волосков, па эпидерме ряда видов заметны выросты, называемые энергенцами К ним относятся известные жгучие «волоски» крапивы, шипы розы, малины, ежевики и др Шипы на плодах многих зонтичных, дурмана, каштана также являются эмергенцами В формировании эмергенцев принимают участие не только клетки эпидермы, но и лежащие под ней слои клеток