

Карты презумптивных зачатков
ПРЕЗУМПТИВНЫЕ ЗАЧАТКИ
( от
лат. praesumptivus — предполагаемый, ожидаемый),
области яйца или раннего зародыша, из
к-рых развиваются те или иные органы. С
помощью методов прижизненного мечения
эмбриональных тканей и прослеживания
дальнейшей судьбы меток составлены
карты П. з. для яиц или ранних зародышей
осн. групп животных
от
лат. praesumptivus — предполагаемый, ожидаемый),
области яйца или раннего зародыша, из
к-рых развиваются те или иные органы. С
помощью методов прижизненного мечения
эмбриональных тканей и прослеживания
дальнейшей судьбы меток составлены
карты П. з. для яиц или ранних зародышей
осн. групп животных
Карта относительного положения и размеров презумптивных зачатков на поверхности ранней гаструлы у земноводных (со спинной стороны): 1 — презумптивный материал энтодермы; 2 — головной энтодермы; 3 — сомитов; 4 — эпидермиса; 5 — нервной пластинки; 6 —- хорды; 7 — боковой мезодермы; 8 — спинная губа бластопора; ан — анимальный полюс; вег — вегетативный полюс.
Осевой комплекс зачатков состоит из:
н
 ервная
пластинка
ервная
пластинкахорда (хордальная пластинка)
лежащие латерально по отношению к хордальной пластинке - зачатки мезодермы.
кишечная энтодерма.
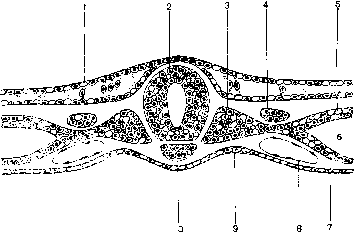
Закладка осевых органов и эмбриональных зачатков у птиц (ориг.):1 - эктодерма- 2 - нервная трубка; 3 – сомит; 4 – нефротом; 5 – париетальный листок спланхнотома; 6 – целом; 7 – висцеральный листок спланхнотома; 8 – кровяной островок; 9 – энтодерма; 10 – хорда.
14 . Вопр Рание этапы развития млеков.
Яйцеклетка - вторично алецитальные (не содержат желтка или имеют незначительное количество желточных включений)
Женские половые клетки, или яйцеклетки, имеют больший размер, чем иные клетки тела, гаплоидный набор хромосом и способны после оплодотворения или партеногенетически развиваться в новый организм.
Х арактерной
особенностью яйцеклетки является также
наличие в ней запасных питательных
веществ в виде желтка, необх для развит
нов организма, наличие особого
поверхностного, или кортикального, слоя
цитоплазмы и специальных оболочек,
покрывающих яйцеклетку. Яйцеклетка в
основном имеет округлую форму, и величина
ее зависит от количества желтка в
цитоплазме. Желток в яйцеклетке находится
в виде пластинок, гранул и состоит из
белков, фосфолипидов, нейтральных жиров.
Ядро яйцеклеток имеет сферическую
форму, содержит одно или множество
ядрышек. Яйцеклетка может иметь до трех
оболочек. Различают первичную,
вторичную
и третичную
оболочки.
Снаружи цитоплазма яйцеклетки покрыта
плазмалеммой, которая образует складки
или мкрвор. Плазмалемму яйца вместе с
веществом, которое располагается между
микроворсинками, называют первичной
5).
Рис.
18. Яйцеклетка млекопитающего:
1
— ядро; 2 —ядрышко; 3
— цитоплазма;
4
— первичная
оболочка; 5 — вторичная оболочка.
Вторичная
оболочка
образована
фолликулярными клетками, которые
располагаются снаружи от первичной
оболочки.. Поскольку отростки фолликулярных
клеток тонкие и прозрачные, то эта часть
вторичной оболочки получила название
светлой
или
блестящей
зоны,
а та часть вторичной оболочки, где
сконцентрированы ядра фолликулярных
клеток, получила название лучистого
венца.Третичная
оболочка
образуется
после выхода яйцеклетки из яичника, при
прохождении ее по яйцеводу, за счет
секреторной деятельности клеток
последнего. Примером третичной оболочки
могут быть студенистая
оболочка яйцеклеток
земноводных, пергаментная
и
скорлуповая
оболочки яйцеклеток
рептилий, белковая,
подскорлуповая и
скорлуповая
оболочки яйцеклеток
птиц.
арактерной
особенностью яйцеклетки является также
наличие в ней запасных питательных
веществ в виде желтка, необх для развит
нов организма, наличие особого
поверхностного, или кортикального, слоя
цитоплазмы и специальных оболочек,
покрывающих яйцеклетку. Яйцеклетка в
основном имеет округлую форму, и величина
ее зависит от количества желтка в
цитоплазме. Желток в яйцеклетке находится
в виде пластинок, гранул и состоит из
белков, фосфолипидов, нейтральных жиров.
Ядро яйцеклеток имеет сферическую
форму, содержит одно или множество
ядрышек. Яйцеклетка может иметь до трех
оболочек. Различают первичную,
вторичную
и третичную
оболочки.
Снаружи цитоплазма яйцеклетки покрыта
плазмалеммой, которая образует складки
или мкрвор. Плазмалемму яйца вместе с
веществом, которое располагается между
микроворсинками, называют первичной
5).
Рис.
18. Яйцеклетка млекопитающего:
1
— ядро; 2 —ядрышко; 3
— цитоплазма;
4
— первичная
оболочка; 5 — вторичная оболочка.
Вторичная
оболочка
образована
фолликулярными клетками, которые
располагаются снаружи от первичной
оболочки.. Поскольку отростки фолликулярных
клеток тонкие и прозрачные, то эта часть
вторичной оболочки получила название
светлой
или
блестящей
зоны,
а та часть вторичной оболочки, где
сконцентрированы ядра фолликулярных
клеток, получила название лучистого
венца.Третичная
оболочка
образуется
после выхода яйцеклетки из яичника, при
прохождении ее по яйцеводу, за счет
секреторной деятельности клеток
последнего. Примером третичной оболочки
могут быть студенистая
оболочка яйцеклеток
земноводных, пергаментная
и
скорлуповая
оболочки яйцеклеток
рептилий, белковая,
подскорлуповая и
скорлуповая
оболочки яйцеклеток
птиц.
Процесс слияния мужской и женской половых клеток, приводящий к образованию зиготы, которая дает начало новому организму, называется 1.оплодотворением. У млеков внутреннее моноспермное, в нач отделах яйцеводов.
осущ. В 2 этапа: I. дистантное (осеменение) взаимодействие, включает в себя 3 механизма: хемотаксис – направленное движение сперматозидов навстречу к яйцеклетке; реотаксис – движение сперматозоидов в половых путях против тока жидкости;
стериотаксис – пособность сперматозоидрв двигатся к более крупному обьекту.
II. контактное взаимодействие: акросомная реакция сперматозоидов- апикальная часть спер-ов, вступает в контакт со студенистой оболочкой яйца. Протеазы акросомы растворяют студенистую оболочку . акр. Вырост несет на себе спецефич белок , а на поверхности яцца имебтся рецепторы узнающие этот белок; активизация яйцеклетки-мембраны обоих клеток сливаются, по образовавшемуся каналу ядро и центриоль проник в яйцо.
2. Дробление — это ряд последовательных митотических делений зиготы, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные, содержащие ядра клетки меньшего размера. В результате дробления образуются клетки, которые называют бластомерами. У млеков дробление полное, неравномерное асинхронное. Бластоциста представляет собой однослойный пузырек, заполненный жидкостью, в котором различают эмбриобласт (из него развивается зародыш) и трофобласт, обеспечивающий питание зародыша.
3.имплантация: 1 фаза – адгезия зародыш связывается с белками внематочного матрикса эпителия. Эндометрия матки. 2 фаза – инвазия зародыша в стенку матки.
4.Гаструляция После того как сформировалась бластула, начинается следующий этап эмбриогенеза — гаструляция (образование зародышевых листков). В результате гаструляции образуется двухслойный, а затем трехслойный зародыш (у большинства животных) — гаструла. Первоначально образуются наружный (эктодерма) и внутренний (энтодерма) слои. Позже между экто- и энтодермой закладывается третий зародышевый листок — мезодермапроходит в 2 этапа: Деламинация). При таком способе гаструляции клеточные перемещения минимальны или совсем отсутствуют, так как происходит расслоение — наружные клетки бластулы преобразуются в эктодерму, а внутренние формируют энтодерму. миграция — «выселение» части клеток бластодермы в полость бластоцеля (высшие позвоночные). Из этих клеток образуется энтодерма.
5. Образование внезародышевых органов: амнион(внезародышевая энтодерма и внезарод мезенхимаАмнион (водная, амниотическая оболочка), представляет собой полый орган (мешок), заполненный жидкостью (околоплодными водами), в которой находится и развивается зародыш. Основная функция амниона — выработка околоплодных вод, которые обеспечивают оптимальную среду для развития зародыша и предохраняют его от высыхания и механических воздействий.); желточный мешок(пупочный, или пуповинный пузырек) — рудиментарное образование, утратившее функцию вместилища питательных веществ. До 7-8-й недели эмбриогенеза основная его функция — кроветворная. Кроме того, в стенке желточного мешка появляются первичные половые клетки — гонобласты, которые мигрируют в него из области первичной полоски. (внезародышевая энтодерма и внезарод мезенхима); АЛЛАНТОИС.Аллантоис представлен внезародышевой энтодермой и внезародышевой мезодермой амниотической ножки. В развитии человека аллантоис не играет существенной роли и остается недоразвитым. Его роль сводится к проведению сосудов из зародыша по амниотической ножке к хориону. Плацента — это основное связующее звено матери и плода, относится к ворсинчатому гемохориальному типу. Развивающийся трофобласт разрушает ткани слизистой оболочки матки и сосуды, формируются лакуны, куда изливается артериальная кровь матери и далее кровь из лакун по венозной системе оттекает из матки. Система мать – плод
Система мать - плод возникает в процессе беременности и включает в себя две подсистемы - организм матери и организм плода, а также плаценту, являющуюся связующим звеном между ними.
Взаимодействие между организмом матери и организмом плода обеспечивается прежде всего нейрогуморальными механизмами. При этом в обеих подсистемах различают следующие механизмы: рецепторные, воспринимающие информацию, регуляторные, осуществляющие ее переработку, и исполнительные.
Рецепторные механизмы организма матери расположены в матке в виде чувствительных нервных окончаний, которые первыми воспринимают информацию о состоянии развивающегося плода.
Регуляторные механизмы организма матери включают отделы центральной нервной системы (височная доля мозга, гипоталамус, мезэнцефальный отдел ретикулярной формации), а также гипоталамоэндокринную систему. Важную регуляторную функцию выполняют гормоны: половые, тироксин, кортикостероиды, инсулин и др. Так, во время беременности происходит усиление активности коры надпочечников матери и повышение выработки кортикостероидов, которые участвуют в регуляции метаболизма плода. В плаценте вырабатывается хорио-нический гонадотропин, стимулирующий образование адрено-кортикотропного гормона гипофиза, который активизирует деятельность коры надпочечников и усиливает секрецию кортикостероидов.
Регуляторные нейроэндокринные аппараты матери обеспечивают сохранение беременности, необходимый уровень функционирования сердца, сосудов, кроветворных органов, печени и оптимальный уровень обмена веществ, газов в зависимости от потребностей плода.
Рецепторные механизмы организма плода воспринимают сигналы об изменениях организма матери или собственного гомеостаза. Они обнаружены в стенках пупочных артерий и вены, в устьях печеночных вен, в коже и кишечнике плода.
Раздражение этих рецепторов приводит к изменению частоты сердцебиения плода, скорости кровотока в его сосудах, влияет на содержание сахара в крови и т. д.
Регуляторные нейрогуморальные механизмы организма плода формируются в процессе развития. Первые двигательные реакции у плода появляются на 2-3-м месяце развития, что свидетельствует о созревании нервных центров. Механизмы, регулирующие газовый гомеостаз, формируются в конце II триместра эмбриогенеза. Начало функционирования центральной эндокринной железы - гипофиза - отмечается на 3-м месяце развития. Синтез кортикостероидов в надпочечниках плода начинается со второй половины беременности и увеличивается с его ростом. У плода усилен синтез инсулина, который необходим для обеспечения его роста, связанного с углеводным и энергетическим обменом. Следует отметить, что у новорожденных, родившихся от матерей, страдающих сахарным диабетом, когда снижена выработка инсулина, наблюдается увеличение массы тела и повышение продукции инсулина в островках поджелудочной железы.
Действие нейрогуморальных регуляторных систем плода направлено на исполнительные механизмы - органы плода, обеспечивающие изменение интенсивности дыхания, сердечно-сосудистой деятельности, мышечной активности и т. д. и определяющие изменение уровня газообмена, обмена веществ, терморегуляции и других функций.
Как уже указывалось, в обеспечении связей в системе мать - плод особо важную роль играет плацента, которая способна не только аккумулировать, но и синтезировать вещества, необходимые для развития плода. Плацента выполняет эндокринные функции, вырабатывая ряд гормонов: прогестерон, эстроген, хорионический гонадотропин, плацентарный лактоген и др. Через плаценту Между матерью и плдом осуществляются гуморальные и нервные связи. Существуют также экстраплацентарные гуморальные связи через плодные оболочки и амниотическую жидкость.
Гуморальный канал связи - самый обширный и информативный. Через него происходит поступление кислорода и углекислого газа, белков, углеводов, витаминов, электролитов, гормонов, антител и др. В норме чужеродные вещества не проникают из организма матери через плаценту. Они могут начать проникать лишь в условиях патологии, когда нарушена барьерная функция плаценты. Важным компонентом гуморальных связей являются иммунологические связи, обеспечивающие поддержание иммунного гомеостаза в системе мать - плод.
Несмотря на то, что организм матери и плода генетически чужеродны по составу белков, иммунологического конфликта обычно не происходит. Это обеспечивается рядом механизмов, среди которых существенное значение имеют: 1 -синтезируемые синцитиотрофобластом белки, тормозящие иммунный ответ материнского организма; 2 - хориональный гонадотропин и плацентарный лактоген, находящиеся в высокий концентрации на поверхности синцитиотрофобласта, принимают участие в угнетении Материнских лимфоцитов; 3-иммуномаскирующее действие гликопротеинов перицеллюлярного фибриноида плацеты, заряженного так же, как и лимфоциты омывающей крови, отрицательно; 4 - протеолитические свойства трофобласта также способствуют инактивации чужеродных белков. В иммунной защите принимают участие и амниотические воды, содержащие антитела, блокирующие антигены А и В, свойственные крови беременной, и не допускают их в кровь плода в случае несовместимой беременности.
Показана определенная взаимосвязь гомологичных органов матери и плода: поражение какого-либо органа матери ведет к нарушению развития одноименного органа плода. В эксперименте на животных установлено, что сыворотка крови животного, у которого удалили часть какого-либо органа, стимулирует пролиферацию в одноименном органе. Однако механизмы этого явления изучены недостаточно.
Вопр 15 ген, реализация генетической информации. Современные представления о структуре гена.
Ген - функциональная единица наследственного материала. Ген (от греч. genos -- род, происхождение) - участок молекулы геномной нуклеиновой кислоты, характеризуемый специфической для него последовательностью нуклеотидов, представляющий единицу функции, отличной от функций д Хранящаяся в ДНК генетическая информация реализуется в процессе биосинтеза белка.
ДНК сосредоточена в ядре клетки, а белки синтезируются в цитоплазме на рибосомах. Для биосинтеза белка необходимо доставить генетическую информацию из ядра клетки к рибосомам. Роль посредника, обеспечивающего передачу генетической информации от ядра клетки к рибосомам, выполняют матричные, или информационные, РНК (мРНК, или иРНК).
Матричные РНК представляют собой полинуклеотидные цепочки с молекулярными массами от 150 тысяч до 5 миллионов дальтон. Они синтезируются в ядре клетки. В ходе биосинтеза мРНК генетическая информация «переписывается» с небольшого участка ДНК, включающего один или несколько генов, на молекулу мРНК. Синтез матричной РНК на значащей нити ДНК получил название транскрипции (лат. "transcriptio " — переписывание).
Процесс транскрипции генетической информации сходен с процессом репликации ДНК. Биосинтез мРНК начицается с расплетания двойной спирали ДНК на небольшом участке.
Свободные рибонуклеозидтрифосфаты с помощью Водородных связей присоединяются к нуклеотидам расплетенного участка ДНК в соответствии с принципом комплементарности
Азотистых оснований.
Образование мРНК происходит путем переноса от рибонуклеозидтрифосфатов остатков рибонуклеотидов к третьему атому углерода рибозы концевого нуклеотида синтезируемой полинуклеотидной цепи. При этом происходит разрыв Макроэргических связей в молекулах рибонуклеозидтрифосфатов с выделением пирофосфата, что обеспечивает процесс транскрипции необходимой энергией. Биосинтез мРНК катализирует фермент РНК-полимераза.
Большую роль в процессе транскрипции играют специальные белки, которые тонко регулируют его ход.
Синтезированная в процессе транскрипции мРНК Поступает из ядра клетки в рибосому — цитоплазматическую серганеллу, по химической природе представляющую собой нукдеопротеид — сложный белок, небелковым компонентом которого является рибонуклеиновая кислота.
РНК. участвующие в построении тела рибосомы («рибонуклеиновая кислота» + гр. «сома» — тело), называют рибосомальными (рРНК). Рибосомы построены из двух субчастиц — большой и малой. В построении каждой из них участвуют большое количество разных белков и различные рРНК. Молекулярная масса рибосомальных РНК колеблется от 55000 до 1600000 дальтон и более. Синтез рРНК, также как и синтез мРНК, происходит в ядре клетки и контролируется ДНК.
Матричная РНК закрепляется в рибосоме. Теперь рибосоме необходимо воспроизвести полученную информацию, записанную в нуклеотидной последовательности мРНК четырехбуквенным «языком» азотистых оснований, на двадцатибуквенном «языке» в виде последовательности аминокислот в полипептидной цепочке синтезируемого белка. Процесс перевода генетической информации с «языка» азотистых оснований на «язык» аминокислот называют трансляцией (лат. "translation" — передача).
Доставку аминокислот к рибосомам обеспечивают транспортные РНК (тРНК). Молекулярные массы тРНК относительно невелики и варьируют в пределах от 17000 до 35000 дальтон. Синтезом тРНК в клетке управляет ДНК.
Процесс биосинтеза белка требует энергетических затрат. Для того чтобы аминокислоты соединились друг с другом пептидной связью, их необходимо активировать. Аминокислоты активируются с участием АТФ и тРНК. Эти реакции катализирует фермент аминоацил-тРНК-синтетаза.
Реакции активирования каждой из протеиногенных аминокислот катализируются своей аминоацил-тРНК-синтетазой.
Эти ферменты позволяют аминокислотам и тРНК безошибочно узнавать друг друга. В результате каждая аминокислота присоединяется к конкретной тРНК. Транспортные РНК называют по присоединяющейся аминокислоте, например: валиновая тРНК, аланиновая тРНК, сериновая тРНК и т. д.
Полинуклеотидные цепочки тРНК имеют пространственную структуру, напоминающую по форме клеверный лист. К одному из концов тРНК присоединяется аминокислота. На другой стороне молекулы тРНК в одной из петель «клеверного листа» имеется триплет нуклеотидов, называемый антикодоном. Этот антикодон комплементарен одному из триплетов мРНК — кодону. Генетический код кодона соответствует аминокислоте, соединенной с тРНК, обладающей комплементарным антикодоном.
Кодоны в зрелой мРНК следуют один за другим непрерывно: они не отделены друг от друга некодирующими участками и не перекрываются.
Аминоацил-тРНК последовательно поступают в рибосомы.
Здесь всякий раз между комплементарными антикодоном тРНК и кодоном мРНК возникают водородные связи. При этом аминогруппа последующей аминокислоты взаимодействует с
Карбоксильной группой предыдущей аминокислоты с образованием пептидной связи.
Синтез любого белка в клетке всегда начинается с N-конца. После образования между аминокислотами пептидной связи рибосома перемещается вдоль цепи мРНК на один кодон. Когда рибосома достигает участка мРНК, содержащего один из трех «бессмысленных» триплетов — УАА, УАГ или УГА, дальнейший синтез полипептидной цепи обрывается. Для этих триплетов в клетке не существует тРНК с комплементарными антикодонами. «Бессмысленные» триплеты располагаются в конце каждого гена и показывают, что синтез данного белка на этом необходимо завершить. Поэтому эти триплеты называют терминирующими (лат. "terminalis" — конечный). По окончании процесса трансляции генетического кода полипептидная цепочка покидает рибосому и формирует свою пространственную структуру, после чего белок приобретает способность к реализации присущей ему биологической функции. Процесс реализации генетической информации в результате транскрипции и трансляции называют экспрессией (лат. "expressio" — выражение) гена.
Биосинтез белка в клетке протекает не на отдельной рибосоме.
Матричная РНК связывается одновременно с несколькими рибосомами, при этом образуется полирибосомальный комплекс. В результате в клетке происходит синтез сразу нескольких одинаковых молекул белка.
ругих генов, и способный изменяться путем мутирования.
Генетический код – это система записи генетической информации в молекуле нуклеиновой кислоты о строении молекулы полипептида, а именно, о количестве, последовательности расположения и типах аминокислот. В одном гене записана информация об одной полипептидной цепочке, т.е. о первичной структуре белка.
Генетический код характеризуется триплетностью, т.е. три нуклеотида, расположенные последовательно в цепочке нуклеиновой кислоты (ДНК или РНК), образуют триплет или кодон (кодовое слово), который кодирует одну аминокислоту и ее местоположение в пептидной цепи. Кодоны различаются последовательностью и типами нуклеотидов (азотистых оснований). Существует 64 типа кодонов, что соответствует количеству возможных сочетаний из 4 (4 типа нуклеотидов, различающихся азотистыми основаниями) по 3 (43). 61 из них – информативные кодоны, они определяют (кодируют) аминокислоты. 3 кодона (в ДНК – АТТ, АТЦ, АЦТ, соответственно в иРНК – УАА, УАГ, УГА) называют стоп-кодонами, они обеспечивают окончание синтеза белковой цепочки. Кодон ТАЦ в ДНК или АУГ в иРНК (кодирует аминокислоту метионин) – стартовый, т.е. стоит первым в гене и с него начинается синтез пептида.
При расшифровке генетического кода оказалось, что большинство аминокислот кодируются несколькими разными кодонами, другими словами, существуют кодоны – синонимы, которые различаются часто только третьими нуклеотидами (азотистыми основаниями). Например, кодоны в ДНК ЦГА, ЦГГ, ЦГТ кодируют аланин, а кодоны ГЦА, ГЦГ, ГЦТ, ГЦЦ, ТЦТ, ТЦЦ – аргинин. Это свойство генетического кода называется вырожденностью или избыточностью.
Вместе с тем было показано, что один кодон кодирует только одну аминокислоту, т.е. в нем может быть записана информация только об одной аминокислоте – иными словами, генетический код однозначен.
Генетический код обладает также неперекрываемостью, это означает, что кодоны располагаются линейно, и один нуклеотид входит в состав только одного кодона; и непрерывностью – кодоны не отделены один от другого, располагаются в цепи нуклеиновой кислоты друг за другом, т.е. расстояние между кодонами соответствует расстоянию между нуклеотидами, а какие-либо сигналы, указывающие на начало или конец кодонов, отсутствуют.
Универсальность генетического кода подразумевает, что генетический код всех организмов характеризуется одинаковыми свойствами (триплетностью, вырожденностью и т.д.); и что смысл кодонов у всех организмов один и тот же (исключение составляют некоторые кодоны митохондрий и бактерий).
У всех прокариотических и эукариотических организмов генетическая информация записана только в одной цепи ДНК, которая называется кодогенной (информативной или значащей) и обозначается знаком "+", вторая цепь не несет генетической информации – некодогенная (неинформативная или незначащая), и обозначается знаком "–