

ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ АВТОНОМНОЕ ОБЩЕОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗОВАНИЯ «НАЦИОНАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ ЯДЕРНЫЙ УНИВЕРСИТЕТ «МИФИ»
ОБНИНСКИЙ ИНСТИТУТ АТОМНОЙ ЭНЕРГЕТИКИ (ИАТЭ) ИНЖЕНЕРНО-ФИЗИЧЕСКИЙ ИНСТИТУТ БИОМЕДИЦИНЫ (ИФИБ) МЕДИЦИНСКИЙ ФАКУЛЬТЕТ
КАФЕДРА МИКРОБИОЛОГИИ, ВИРУСОЛОГИИ, ИММУНОЛОГИИ
дисциплина «Клиническая иммунология и вирусология»
Доклад на тему
«Иммунитет при вирусных инфекциях»
Студент 6 курса ЛД2-С13Б группы Тюлякова Эльвира Николаевна
Преподаватель: зав. кафедрой Микробиологии, вирусологии, иммунологии
2019 |
Колесникова Светлана Геннадьевна |
|

• Механизмы противовирусного иммунитета имеют особенности, определяемые природой вирусов, паразитизмом на уровне молекулярных и субмолекулярных структур. Внеклеточный вирус не проявляет своего патогенного действия, а иммунные реакции против внеклеточных вирионов сходны с
реакциями на другие
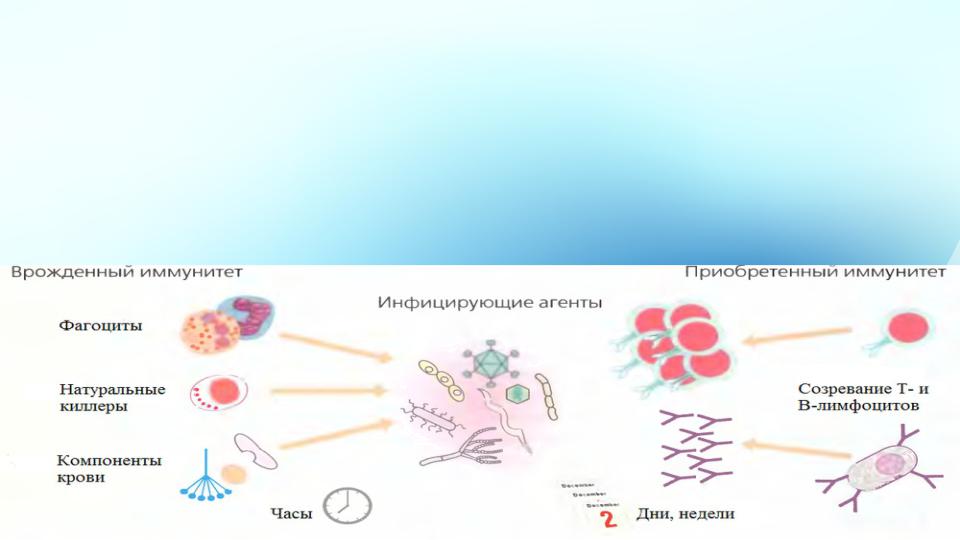
•Ранняя стадия вирусной инфекции, как правило, состоит в противоборстве вируса с защитными системами организма- хозяина. В случае преодоления вирусами кожного покрова и слизистых оболочек организма в действие вступают механизмы экстренной неспецифической защиты (врожденного иммунитета ): макрофаги , интерфероны , НК- клетки (нормальные киллеры) .

Неспецифический
иммунный ответ
•Запуск иммунных процессов, связанных с активацией врожденного иммунитета, зависит главным образом от распознавания вирусных нуклеиновых кислот (в случае ретровирусов - двуспиральной РНК) патогенраспознающими рецепторами, локализованными не на поверхности, а в цитоплазматических гранулах клеток врожденного иммунитета (TLR-3, TLR-7, TLR-8, TLR-9).
•Оболочка вирусов содержит белки, распознаваемые в большей степени В-клетками и продуцируемыми ими антителами. В вирусе гриппа это гемагглютинин и нейраминидаза. Серотип вируса, определяющий специфичность образуемых антител, выражают формулой, содержащей обозначения типа этих молекул (например, H2N5).

• Инфицированные клетки гибнут вследствие цитопатогенного действия вируса. Их фрагменты поглощаются макрофагами и дендритными клетками (первые поглощают как корпускулярные фрагменты, так и растворимые компоненты, вторые - преимущественно растворимые молекулы).
• Вирусные компоненты, ДНК, одно- и двуспиральная РНК распознаются TLR внутри фаголизосом и сигнализируют о появлении чужеродных агентов - носителей PAMP. Индуцируемые при этом внутриклеточные сигналы приводят к реализации двух главных эффектов - активации (через транскрипционный фактор NF-kB) генов провоспалительных факторов и индукции синтеза интерферонов типа I (a и β).
• Провоспалительные сигналы реализуется особенно интенсивно в
макрофагах. Они активируются и инициируют развитие локального воспаления, сопровождающегося секрецией провоспалительных
цитокинов.

•Продукция и секреция цитокинов ИФН α/β, ИЛ-1, ФНО, ИЛ-6, 8 относится к самым ранним событиям, сопутствующим взаимодействию микроорганизмов с макрофагами; этот ранний неспецифический ответ на инфекцию важен по нескольким причинам:
•он развивается очень быстро, поскольку не связан с необходимостью накопления клона клеток, отвечающих на конкретный антиген;
•вместе с тем ранний цитокиновый ответ влияет на последующий специфический иммунный ответ

Интерфероны типа I наиболее интенсивно синтезируют плазмоцитоидные дендритные клетки, в меньшей степени - макрофаги. Интерфероны типа I служат факторами противовирусной защиты, вызывая деградацию вирусной РНК и препятствуя репликации вирусов (защитный фактор 1).
• Интерферон активирует макрофаги и ЕК-клетки, которые затем синтезируют гамма-ИФН, ИЛ-1, 2, 4, 6, ФНО, в результате макрофаги и ЕК-клетки приобретают способность лизировать вирус- инфицированные клетки

•ИФН-гамма индуцирует и стимулирует продукцию провоспалительных монокинов (ФНО, ИЛ-1, 6), экспрессию на мембранах макрофагов, антигенов МНС II; ИФН-гамма резко усиливает антимикробную и противовоспалительную активность за счет повышения продукции клетками супероксидных радикалов, а усиление иммунного фагоцитоза и антител-опосредованной цитотоксичности макрофагов под влиянием ИФН-гамма связаны с усилением экспрессии Fc-рецепторов для иммуноглобулина G.

Этой системе (интерфероновой) принадлежит главная роль в выздоровлении от вирусных инфекций, выражающаяся в защите соседних с зараженными клеток от инвазии их вирусами.
При встрече организма с вирусной инфекцией именно продукция интерферона (растворимого фактора, вырабатываемого вирус- инфицированными клетками, способного индуцировать антивирусный статус в неинфицированных клетках) является наиболее быстрой ответной реакцией на заражение (Hall М. et аl, 1992). Интерферон формирует защитный барьер на пути вирусов намного раньше специфических защитных реакций иммунитета, стимулируя клеточную резистентность, - делая клетки непригодными для размножения вирусов.

Главными эффекторами противовирусного иммунитета на этом этапе служат естественные киллеры, активируемые при распознавании стрессорных молекул инфицированных клеток и дополнительно стимулируемые цитокинами, которые секретируют NKT-клетки.
На поверхности инфицированной клетки экспрессируются стрессорные белки (MICA, MICB, ULBP), сигнализирующие о
нарушении физиологического состояния клетки. Эти белки распознаются NK-клетками. Активированные NK-клетки
осуществляют цитолиз клеток-мишеней, выступая в качестве фактора противовирусной защиты,
функционирующего в рамках врожденного иммунитета (защитный фактор 2).