

2.1 Структура Ту-елементів
[Lee F.W.F. et al. // Appl Microbiol Biotechnol – 1997 – vol.48 – P. 339-345.]
Ту-елементи S. cerevisiae (“transposons of yeast”)– сімейство мобільних генентичних елементів геному, які присутні у 35-ти копіях в гаплоїдному геномі дріжджів; на кінцях мають довгі термінальні повтори по 340 пар основ (англ. long terminal repeats - LTR), також відомі як δ-послідовності.

Рис 2. Структура Ту1-елемента дріжджів [Modern Genetic Analysis.
Griffiths AJF, Gelbart WM, Miller JH, et al.New York: 1999.]
δ-послідовності – це довгі кінцеві повтори нуклеотидів, що входять до складу ретротранспозонів Ty1 i Ty2, що вбудовуються в геномну ДНК дріжджів. Копії цих нуклеотидних послідовностей залишаються у вихідній ділянці молекули ДНК після того, як відбулася транспозиція Ty-елемента в іншу ділянку, тому в геномі дріжджів присутня також велика кількість ізольованих δ-послідовностей. Загалом налічується близько 425 таких послідовностей у геномній ДНК S. cerevisiae
На даний час відомо 5 основних типів Ту-елементів: Ty1 (3), Ty2 (22), Ty3 (10), Ty4 (11) і Ty5 (3), які розрізняються за довжиною, розміщенням сайтів рестриктаз, кількістю копій у гаплоїдному геномі, а також за сайтами інсерції. Ту-елементи містять гени складаються з двох генів: TYA1 та TYB1, що відповідають генам gag і pol ретровірусів [Curcio M.J., Garfinkel D.J. (1990) Genetics,88, 136 – 140.]..
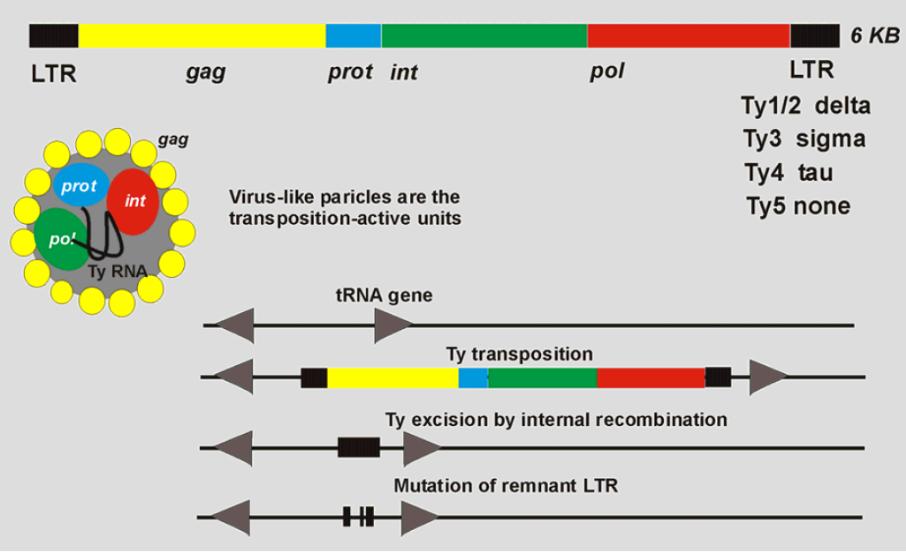
Рис 3. Ту-елементи дріжджів [TY ELEMENTS OF THE YEAST SACCHAROMYCES CEREVISIAE O. Krastanova1, M. Hadzhitodorov2, M. Pesheva1 Sofia University "St. Kl. Ohridski", Faculty of Biology, Microbial Genetics Laboratory,Sofia, Bulgaria1]

Рис 4. Класифікація Ту-елементів S. cerevisiae.
За своєю структурою Ту-елементи подібні до довгих термінальних повторів еукаріотичних ретровірусів, оскільки вони позбавлені деяких функцій даних повторів і розглядаються як примітивні ретровіруси [The EMBO Journal Vol.2 No.4 pp.583-591, 1983 Delta sequences in the 5' non-coding region of yeast tRNA genesJurg Gafner, Eddy M.De Robertis1 and Peter Philippsen]. Саме через ці подібності з ретровірусами Ту-елементи також мають назву ретротранспозони.
δ-послідовності дріжджів, так як і Ту-елементи загалом, показують дивергенцію у послідовностях. Термінальні δ-послідовності є прямим повторами, на відміну від мобільних елементів бактерій, які містять інвертовані повтори (англ. inverted repeats – IR). Однак, як і прокарітоичні як і прокарітотичні транспозони, Ту-елементи генерують повторювану послідовність ДНК-мішені (5 тис.пар основ), а також спричиняють мутації, випадково вбудовуючись у сайти геному. Однією з основних ознак ретротранспозонів дріжджів є їх випадкова мультикопійна інтеграція у різні сайти геному клітини-хазяїна
2.2 Транспозиція Ту-елементів
Ту-мРНК транскрибується і піддається процесингу у ядрі і згодом транспортується у цитоплазму, де слугує матрицею для трансляції Gag та Gag-Pol білків [Garfinkel D.J. (1992) “Retroviral” Retroelementsin Microorganisms”, Plenum press, N.Y., 107– 136.]. Ту-елементи S. cerevisiae продукують вірусоподібні частинки (англ. virus-like particles, VLPs), в яких під час процесу збірки відбувається пакування РНК, з якої шляхом зворотної транскрипції відбувається синтез повнорозмірної кДНК. На фінальному етапі транспозиції кДНК інтегрується у новий сайт геному і цикл може відбуватися повторно [Lodish,Berk,Zipursky, Baltimore, Darnell ‘Molecular cell biology’. W.H Freeman and company, Section 9.3 Mobile DNA]
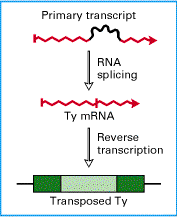
Рис 5. Транспозиція і зворотня транскрипція Ту-елемента.
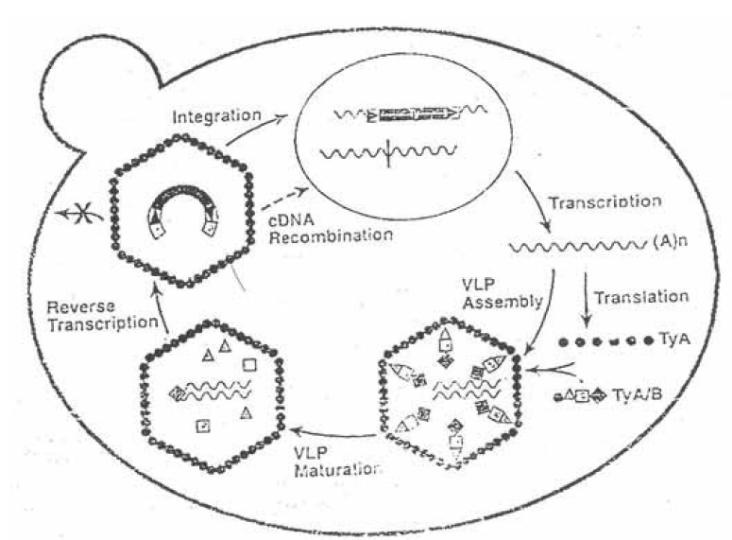
Рис 6. Життєвий цикл ретротранспозона Saccharomyces cerevisiae:
1 – Транскрипція з Ту-елемента з утворенням TyA/B мРНК;
2 – Пакування TyA/B мРНК у вірусоподібні частинки;
3 – Дозрівання вірусоподібних частинок
4 – Зворотня транскрипція з мРНК з утворенням кДНК;
5 – Інтеграція кДНК у геном шляхом гомологічної рекомбінації.