

Функции цикла Кребса:
1.Водороддонорная функция. Цикл Кребса поставляет субстраты для дыхательной цепи (НАД-зависимые субстраты: изоцитрат, -кетоглутарат, малат; ФАД-зависимый субстрат – сукцинат). 2.Катаболическая функция. В ходе ЦТК окисляются до конечных продуктов обмена ацетильные остатки, образовавшиеся из топливных молекул (глюкоза, жирные кислоты, глицерол, аминокислоты). 3.Анаболическая функция. Субстраты ЦТК являются основой для синтеза многих молекул (кетокислоты — α-кетоглутарат и ЩУК — могут превращаться в аминокислоты глу и асп; ЩУК может превращаться в глюкозу, сукцинил-КоА используется на синтез гема). 4.Анаплеротическая функция. Цикл не прерывается благодаря реакциям анаплероза (пополнения) фонда его субстратов. Важнейшей анаплеротической реакцией является образование ЩУК (молекулы, запускающей цикл) путем карбоксилирования ПВК. 5.Энергетическая функция. На уровне сукцинил-КоА происходит субстратное фосфорилирование с образованием 1 молекулы макроэрга. Помимо этого, 4 дегидрогеназные реакции в цикле Кребса создают мощный поток электронов, богатых энергией. Эти электроны поступают в дыхательную цепь внутренней мембраны митохондрий. Конечным акцептором электронов является кислород. При последовательном переносе электронов на кислород выделяется энергия, достаточная для образования 9 молекул АТФ путем окислительного фосфорилирования. Примечание: более понятной эта цифра станет после того, как мы познакомимся с работой дыхательной цепи и с ферментом, синтезирующим АТФ.
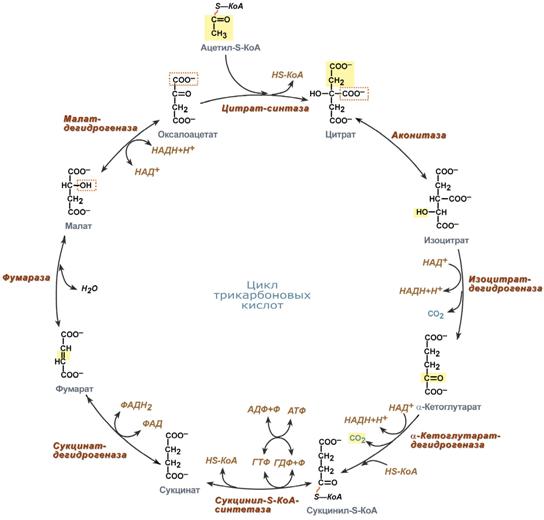
41.
Тканевой липолиз. Он происходит в жировой ткани постоянно. Гидролиз тканевых жиров осуществляет высокоспецифичный фермент гормогочувствительная липаза (ТАГ-липаза). Оно гидролизует ТАГ до глицерина и свободных жирных кислот.
Глицерин не утилизируется в клетках жировой ткани, поэтому он выходит из адипоцитов и поступает в плазму, а из кровотока – в печень и далее в почки. В клетках печени и почек активен фермент глицерокиназа, поэтому только эти органы используют глицерин.
Свободные жирные кислоты в жировой ткани могут вновь активироваться и использоваться для синтеза ТАГ.
Таким образом, в жтровой ткани с одинаковой скоростью идут 2 противоположных процесса: распад и синтез ТАГ.
Если скорость липолиза превышает скорость синтеза жиров(биополимеры), то свободные жирные кислоты поступают в плазму, где связываются с сывороточными альбуминами, которые переносят их ко всем органам (важный источник энергии): печень, скелетная и сердечная мышцы. Следовательно, транспортной формой ТАГ от жирового депо к другим органам являются альбумины плазмы.
Фермент гормоночувствительная (ГЧ) липаза (ТАГ-липаза) активируется адреналином, норадреналином, глюкагоном, АКТГ. Эти гормоны обладают непрямым механизмом действия и активируют гормоночувствительную липазу через вторичных посредников: ц-ФМФ, ионов Са, диацилглицерина (ДАГ) и инозитолтрифосфата (ИФЗ). Инсулин-их антаганист, т.к. ингибирует ГЧ липазу.
При сахарном диабете в отсутствии инсулина в крови резко возрастает содержание свободных высших жирных кислот, которые клетки используют как источник энергии, вместо глюкозы.
Триглицериды. Основные функции:
Увеличивают энергопродукцию.
Повышают выносливость.
Сберегают гликоген мышц.
Уменьшают количество жира в теле.
Триглицериды со средней цепью (ТСЦ) — соединения с атомами углерода, которые дают выход. Они водорастворимые и легко абсорбируются в систему печени через воротную вену. Их применяют в медицинской практике в течение многих лет для лечения нарушений всасывания жира, дефектов во всасывании липидов в результате кишечных реакций и недостаточного транспорта липидов. Общепринято, что повышение окисления свободных жирных кислот снижает утилизацию мышечного гликогена. Поскольку ТСЦ окисляются так же быстро, как и глюкоза, теоретически они могут улучшить спортивные показатели путем сбережения мышечного гликогена во время выполнения упражнений. Основываясь на этой теории, производители заявляют, что ТСЦ усиливают энергопродукцию, повышают выносливость, снижают жировую и увеличивают тощую массу.
37.
Спиртовым брожением называется процесс расщепления сахара микроорганизмами с образованием этилового спирта и углекислого газа.
С6Н12О6 – 2СН3СН2ОН+2СО2
Возбудителями спиртового брожения являются дрожи сахаромицеты, некоторые мицеальные грибы. Даже растения и грибы в анаэробных условиях способны накапливать этиловый спирт.
Процесс проходит 2 стадии:
1. Окислительная – превращение глюкозы до пировиноградной кислоты (пируват) и отнятие двух пар водорода.
С6Н12О6–2СН3СОСООН= НАД (кофермент) Н2О
2. Далее пируват декарбоксилируется пируваткарбоксилазой при участии тиаминпирофрсфата до ацетальдегида, а затем ацетальдегид восстанавливается алкогольдегидрогеназой в этанол при участии кофермента НАД.
Во второй стадии – восстановительный кофермент НАД Н2 передает водород конечному акцептору.
Химизм молочнокислого брожения.
Процесс превращения глюкозы до пировиноградной кислоты у гомоферментатавных молочнокислых бактерий протекает как и у дрожжей при спиртовом брожении. Далее ввиду отсутствия у этих бактерий пируватдекарбоксилазы, пировиноградная кислота не подвергается расщеплению, она является в этом брожении конечным акцептором водорода. Пировиноградная кислота вступает во взаимодействие с восстановленным НАД Н2 (кофермент) – образуется молочная кислота.брожение бактерия микроорганизм спиртовой
СНзСОСООН + НАД Н2 – СНзСНОНСООН
УСЛОВИЯ СПИРТОВОГО БРОЖЕНИЯ: На развитие дрожжей и ход брожения влияют: химический состав сбраживаемой среды, концентрация и кислотность среды, содержание спирта, температура, наличие посторонних микроорганизмов. Большинство дрожжей способны сбраживать моносахариды, а из дисахаридов-сахарозу и мальтозу. Дрожжи не могут сбраживать крахмал, так как они не образуют амилолитических ферментов. Наиболее благоприятная концентрация сахара- от 10 до 15%. При повышении концентрации сахара энергия брожения снижается, а при 30-35% сахара брожение прекращается. Хорошим источником азота для большинства дрожжей являются аммонийные соли, но дрожжи могут использовать также аминокислоты и пептиды. Нормальное брожение протекает в кислой среде, при рН 4-5. В щелочной среде в результате брожения образуется глицерин. Наибольшая скорость брожения при температуре 30 С. При температуре 45-50оС брожение прекращается в результате гибели клеток дрожжей. Снижение температуры замедляет ход брожения, но полностью оно не прекращается даже при температуре ниже ОоС. С энергетической точки зрения брожение — процесс малоэффективный. Так, если при окислении 1 граммолекулы глюкозы до СО2 и Н2О в процессе аэробного дыхания синтезируется 36 моль АТФ, то в процессе спиртового брожения — всего 2 моль АТФ. Дрожжи могут переключать один тип обмена веществ (аэробный) на другой (анаэробный). По характеру брожения дрожжи подразделяют на верховые и низовые. Брожение, вызываемое верховыми дрожжими, протекает быстро и бурно при температуре 20-28 С. На поверхности бродящей жидкости образуется много пены и под действием выделяющегося углекислого газа дрожжи выносятся в верхние слои субстрата. По окончании брожения дрожжи оседают на дно бродильных сосудов рыхлым слоем. Брожение, которое вызывают низовые дрожжи протекает медленно при темературе 5-10 С. Газ выделяется постепенно, пены образуется меньше, дрожжи быстро оседают на дно бродильных емкостей. Этиловый спирт, образующийся в процессе брожения, неблагоприятно влияет на дрожжи. Накопление дрожжами спирта в концентрации 2-5% действует на них угнетающе. В большинстве случаев брожение прекращается при накоплении дрожжами 12-14% (объемных) спирта. В настоящее время выделены расы дрожжей, устойчивые к накоплению даже 20% спирта.
ПРАКТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ СПИРТОВОГО БРОЖЕНИЯ: Процесс спиртового брожения лежит в основе получения этилового спирта, кормовых и пищевых дрожжей, пивоварения, хлебопечения, производства глицерина. Совместно с молочнокислым брожением используется при получении кисломолочных продуктов (кумуса, кефира). Для получения этилового спирта используют разное сырье трех основных групп: содержащее сахар (сахарная свекла, кормовая патока, или меласса, сахарный тростник, фруктовые соки); содержащее крахмал (картофель, земляная груша, кукуруза, ячмень, овес, рожь, пшеница); содержащее целлюлозу (древесина и сульфитные щелока). Сырье используют в зависимости от хозяйственных возможностей; оно должно быть дешевым и в достаточном количестве. Крахмалсодержащее сырье разваривают и подвергают осахариванию. Источником амилолитических ферментов служит солодовое молоко, изготовляемое из проросших зерен ячменя, или ферментный препарат из грибов рода Aspergillus. Вносят и дополни:льные источники питания. Это делается всегда по рецепту сред для каждого данного производства. В полученное сусло вносят дрожжи, чаще всего применяют расы Saccharomyces cerevisiae, которые быстро размножаются, с устойчивы, обладают высокой энергией брожения. Есть и промышленно важные расы дрожжей. По окончании брожения дрожжи отделяют от сброженных заторов, а спирт отгоняют на специальных перегонных аппарата. Получается спирт-сырец и остается отход производства — барда которую используют для получения кормовых дрожжей. Отработанные дрожжи тоже используются в виде жидких и сухих кормовых дрожжей. Спирт-сырец используют как для технических целей, так и для дальнейшей очистки — ректификации.
35.
ГЛЮКОГЕНЕЗ – процесс образования организмами глюкозы из соединений, отличных от углеводов. Обычно рассматривается при росте микроорганизмов на низкомолекулярных соединениях (напр., органических кислотах) в качестве единственного источника углерода. Г. реализуется путем обращения большинства стадий гликолиза.
Химизм - Химическая природа какого-л. вещества, явления, процесса.
Малейшее неравновесие в сложном химизме крови — и животное страдает. Герцен, Письма об изучении природы.
Далее мы увидим, что своим химизмом растение воздействует и на неорганическую природу.
Регуляция - Упорядочение, нормализация.
49.
В настоящее время экспериментально обосновано существование четырех главных этапов распада молекулуглеводов, белков и жиров, которые интегрируют образование энергии из основных пищевых источников. На II этапе мономерные молекулы (гексозы, глицерин, жирные кислоты и аминокислоты) подвергаются дальнейшему распаду, в процессе которого образуются богатые энергией фосфатные соединения и ацетил-КоА. В частности, при гликолизе гексозы расщепляются до пировиноград-ной кислоты и далее до ацетил-КоА. Этот процесс сопровождается образованием ограниченного числа богатых энергией фосфатных связей путем субстратного фосфорилирования. На этом этапе высшие жирные кислоты аналогично распадаются до ацетил-КоА, в то время как глицерин окисляется по гликолитическому пути до пировиноградной кислоты и далее до ацетил-КоА. Для аминокислот ситуация на II этапе несколько отлична. При преимущественном использовании аминокислот в качестве источника энергии (при дефиците углеводов или при сахарном диабете) некоторые из них непосредственно превращаются в метаболиты лимоннокислого цикла (глутамат, аспартат), другие – опосредованно через глутамат (пролин, гистидин, аргинин), третьи – в пируват и далее в ацетил-КоА (аланин, серин, глицин, цистеин). Наконец, ряд аминокислот, в частности лейцин, изо-лейцин, расщепляется до ацетил-КоА, а из фенилаланина и тирозина, помимо ацетил-КоА, образуется оксалоацетат через фумаровую кислоту. Как видно, II этап можно назвать этапом образования ацетил-КоА, являющегося по существу единым (общим) промежуточным продуктом катаболизма основных пищевых веществ в клетках.
На III этапе ацетил-КоА (и некоторые другие метаболиты, например α-кетоглутарат, оксалоацетат) подвергаются окислению («сгоранию») в цикле ди- и трикарбоновых кислот Кребса. Окисление сопровождается образованием восстановленных форм НАДН + Н+ и ФАДН2.
В организмечеловека, как и в живой природе вообще, не существует самостоятельного обмена белков, жиров, углеводови нуклеиновых кислот. Все превращения объединены в целостный процесс метаболизма, подчиняющийся диалектическим закономерностям взаимозависимости и взаимообусловленности, допускающий также взаимопревращения между отдельными классами органических веществ. Подобные взаимопревращения диктуются физиологическими потребностями организма, а также целесообразностью замены одних классов органических веществ другими в условиях блокирования какого-либо процесса при патологии.
Еще Кребс и Корнберг отмечали, что, несмотря на огромное разнообразие пищевых веществ (белки, жиры,углеводы), число химических реакций, обеспечивающих их превращения (распад) и образование энергии, «удивительно мало». Эти закономерности свойственны как организму животных и человека, так и микроорганизмам и растениям.
40.
Переваривание липидов в желудочно-кишечном тракте – сложный биохимический процесс. Для создания физиологических условий гидролиза липидов в кишечнике необходимо полноценное функционирование нескольких органов: печени, поджелудочной железы, тонкого кишечника. Следовательно, переваривание и всасывание липидов в кишечнике- это результат интеграции работы вышеперечисленных органов. Дефект участия этих органов в усвоении пищевых липидов организмом приводит к различным заболеваниям, нарушению обмена жиров. Так, снижение синтеза желчных кислот в печени, патология желчеобразования и желчевыделения приводит к развитию стеатореи (жировой стул), создает препятствие для всасывания продуктов гидролиза жиров, способствует развитию гиповитаминоза группы жирорастворимых витаминов.
Всасываться в клетки могут все продукты переваривания, а в очень небольшой мере — и нерасщепленные жиры. Однако большая часть триацилглицеринов распадается до р-моноацилглицеринов, на долю которых приходится примерно 3/4 всех всасывающихся продуктов. Продукты переваривания жиров вместе с желчными кислотами и фосфолипидами желчи образуют смешанные мицеллы, и затем все компоненты мицелл проникают в клетки слизистой кишечника.
Желчные кислоты затем поступают в кровь, а с ней — в печень и повторно участвуют в образовании желчи. Часть желчных кислот не всасывается и выводится с калом (0,2-0,5 г в сутки). Глицерин как водорастворимое вещество всасывается без участия желчи. При нарушении желчеобразования или выделения желчи (например, вследствие закупорки желчного протока желчным камнем, опухолью) условия переваривания жиров и всасывания продуктов гидролиза ухудшаются, и значительная их часть выводится с калом (стеаторея). Жирорастворимые витамины при этом также не всасываются, что приводит к развитию гиповитаминоза.Часть углеводов, поступающих с пищей, превращается в организме в жиры, особенно если количество углеводов превышает необходимое для возобновления запасов гликогена в печени и мышцах. Схема этого превращения представлена на рис. 10.15. Глюкоза служит источником ацетил-КоА, из которого синтезируются жирные кислоты. Необходимый для восстановительных реакций НАДФН поставляется за счет окисления глюкозы в пентозофосфатном пути, а также за счет дегидрирования яблочной кислоты НАДФ-зависимой малатдегидрогеназой. Глице-рол-3-фосфат получается путем восстановления диоксиацетонфосфата — промежуточного продукта гликолиза (рис. 10.16). Таким образом, из глюкозы образуется все, что необходимо для синтеза жиров. Синтез триацилглицеринов из глицерол-3-фосфата и ацил-КоА представлен на рис. 10.17. Синтез жиров из углеводов наиболее активно происходит в печени, жировой ткани и лактирующих молочных железах.Поскольку жиры и другие липиды нерастворимы или очень малорастворимы в воде и в жидкостях организма, необходимы специальные механизмы для транспорта этих веществ кровью. Транспорт осуществляется в составе особых частиц — липопроте-инов. Липопротеины — многомолекулярные структуры. Они представляют собой сферические частицы, поверхностная часть которых образована монослоем ориентированных фосфолипидов и белками (аполипопротеинами). Фосфолипиды гидрофильными концами образуют наружную поверхность, а гидрофобные концы «растворены» в липидной фазе внутри частиц (рис. 10.18). Эта внутренняя липидная фаза содержит в основном триацилглицерины и эфи-ры холестерина. В крови содержится несколько форм липопротеинов; основные из них — хиломикроны, липопротеины очень низкой плотности (ЛОНП), липопротеины низкой плотности (ЛНП) и липопротеины высокой плотности (ЛВП). Липопротеины различаются по составу и содержанию липидов и белков. Приведенные в таблице величины имеют лишь ориентировочный характер, поскольку в процессе функционирования липопротеинов их состав непрерывно изменяется.
44.
На II этапе образования энергии мономерные молекулы (гексозы, глицерин, жирные кислоты и аминокислоты) подвергаются дальнейшему распаду, в процессе которого образуются богатые энергией фосфатные соединения и ацетил-КоА. В частности, при гликолизе гексозы расщепляются до пировиноград-ной кислоты и далее до ацетил-КоА. Этот процесс сопровождается образованием ограниченного числа богатых энергией фосфатных связей путем субстратного фосфорилирования. На этом этапе высшие жирные кислоты аналогично распадаются до ацетил-КоА, в то время как глицерин окисляется по гликолитическому пути до пировиноградной кислоты и далее до ацетил-КоА. Для аминокислот ситуация на II этапе несколько отлична. При преимущественном использовании аминокислот в качестве источника энергии (при дефиците углеводов или при сахарном диабете) некоторые из них непосредственно превращаются в метаболиты лимоннокислого цикла (глутамат, аспартат), другие – опосредованно через глутамат (пролин, гистидин, аргинин), третьи – в пируват и далее в ацетил-КоА (аланин, серин, глицин, цистеин). Наконец, ряд аминокислот, в частности лейцин, изо-лейцин, расщепляется до ацетил-КоА, а из фенилаланина и тирозина, помимо ацетил-КоА, образуется оксалоацетат через фумаровую кислоту. Как видно, II этап можно назвать этапом образования ацетил-КоА, являющегося по существу единым (общим) промежуточным продуктом катаболизма основных пищевых веществ в клетках.
На III этапе ацетил-КоА (и некоторые другие метаболиты, например α-кетоглутарат, оксалоацетат) подвергаются окислению («сгоранию») в цикле ди- и трикарбоновых кислот Кребса. Окисление сопровождается образованием восстановленных форм НАДН + Н+ и ФАДН2.
В организмечеловека, как и в живой природе вообще, не существует самостоятельного обмена белков, жиров, углеводови нуклеиновых кислот. Все превращения объединены в целостный процесс метаболизма, подчиняющийся диалектическим закономерностям взаимозависимости и взаимообусловленности, допускающий также взаимопревращения между отдельными классами органических веществ. Подобные взаимопревращения диктуются физиологическими потребностями организма, а также целесообразностью замены одних классов органических веществ другими в условиях блокирования какого-либо процесса при патологии.
Еще Кребс и Корнберг отмечали, что, несмотря на огромное разнообразие пищевых веществ (белки, жиры,углеводы), число химических реакций, обеспечивающих их превращения (распад) и образование энергии, «удивительно мало». Эти закономерности свойственны как организму животных и человека, так и микроорганизмам и растениям.