

Лекция №10 механизмы образования условного рефлекса
Функциональные основы замыкания временной связи
Стадии и механизм образования условных рефлексов
Доминантный очаг
Торможение условных рефлексов
1
Постоянную («жесткую») связь с агентами внешней среды и деятельностью организма обеспечивают безусловные рефлексы, а временную («пластичную») — условные рефлексы. Условные рефлексы способны к долговременному сохранению в памяти, а также обладают свойством «изменчивости» (перестройки) в зависимости от среды, ситуации лежит в основе индивидуального приспособления, накопления индивидуального опыта, научения, «знаний».
Все условные рефлексы были разделены на классические и инструментальные. В случае выработки классического условного рефлекса через определенные интервалы после условного раздражителя предъявляется какой-либо подкрепляющий (безусловный) стимул. Подкрепление следует за условным стимулом независимо от того, будет ли реакция животного или ее не будет.
В период упрочения связи между условным стимулом и безусловным подкреплением проба изолированного действия условного стимула вызывает условную реакцию. Условный ответ полностью или частично имитирует безусловную реакцию. В павловской школе индикатором формирования условного рефлекса служила, как известно, величина секреции околоушной слюнной железы собаки. После нескольких сочетаний условного и безусловного раздражителей, например звука и пищи, предъявление условного раздражителя (звука) вызывало слюноотделение. В процедуре сочетания условного и безусловного раздражителей безусловный раздражитель (пища) выступает в роли подкрепления. Если условный раздражитель повторно применять без подкрепления, наблюдается угашение, т.е. условно-рефлекторный ответ постепенно уменьшается и в конечном итоге прекращается. Однако после перерыва вновь восстанавливается.
В инструментальных условных рефлексах значительную роль выполняет внутренняя активность животного. Инструментальные рефлексы – это рефлексы, в которых непременным условием является осуществление тех или иных двигательных реакций (как для получения вознаграждения, так и для избавления от болевого стимула), где движение всегда имеет сигнальное значение. Из определений следует, что инструментальный рефлекс характеризуется активным поведением животного, направленным на получение биологически полезного результата. Этот полезный результат и является подкреплением в инструментальных рефлексах.
Физиологическую основу условного рефлекса составляет процесс замыкания временной связи.
Временная (условная) связь – это совокупность нейрофизиологических, биохимических и ультраструктурных изменений мозга, возникающих в процессе сочетания условного и безусловного раздражителей и формирующих определенные взаимоотношения между различными мозговыми образованиями. Механизм памяти фиксирует эти взаимоотношения, обеспечивая их удержание и воспроизведение.
И.П. Павлов считал, что замыкание временных связей происходит в коре больших полушарий головного мозга между тем пунктом, который воспринимает условный раздражитель, и корковым представительством безусловного рефлекса. Каждый условный сигнал поступает в корковый конец анализатора, в проекционную зону, соответствующую модальности стимула. Каждый безусловный раздражитель, центр которого расположен в подкорковых структурах, имеет свое представительство в коре больших полушарий мозга. Изучая безусловные рефлексы нормальных и декортицированных животных, было установлено, что центральная часть дуги безусловного рефлекса не однолинейна. Она проходит не через один уровень мозга, а через многоуровневую структуру, т.е. центральная часть дуги безусловного рефлекса состоит из многих ветвей, которые проходят через различные уровни ЦНС, спинной мозг, продолговатый мозг, стволовые отделы и т. д. (рис.). Наивысшая часть дуги проходит через кору большого мозга, является корковым представительством данного безусловного рефлекса.

Схема дуги безусловного рефлекса (по Э.Л. Асратяну, 1974). I-V - ветви центральной части дуги на различных уровнях мозга; А - афферентная клетка, Э - эфферентная клетка
Условные рефлексы образуются лишь при большей физиологической силе безусловного раздражителя по сравнению с силой условного. Важнейшим фактором в выработке условной связи является совпадение безусловного подкрепления с доминирующей потребностью организма. Увеличение интенсивности подкрепления обычно повышает эффективность научения (скорость выработки условного рефлекса и его прочность). Эффективность выработки условно-рефлекторного научения определяется как отношение числа условных (правильных) ответов к общему числу предъявлений условного стимула.
Обычно образование условного рефлекса происходит при совпадении сочетаемых раздражителей либо опережении сигнальным раздражителем безусловного подкрепления. Для большинства условных рефлексов наиболее эффективно 0,5-секундное опережение условным стимулом безусловного.
Условные рефлексы по показателю временных соотношений между ассоциируемыми раздражителями делят на две группы:
наличные – в случае совпадения во времени условного сигнала и подкрепления,
следовые условные рефлексы, когда подкрепление предъявляется лишь после окончания условного раздражителя.
Наличные рефлексы в свою очередь по величине интервала между включением раздражителей делят на несколько видов - совпадающие, отставленные и запаздывающие.
При совпадающем условном рефлексе подкрепление сразу присоединяется к сигнальному раздражителю (не позднее 1-3 с), при отставленном — в период до 30 с, а в случае запаздывающего рефлекса изолированное действие условного стимула продолжается 1-3 мин.
Следовые условные рефлексы образуются тогда, когда подкрепление следует уже после окончания действия условного стимула. Иными словами, подкрепление сочетается с остаточными «следами нервного стимула».
Условные рефлексы на время — особая разновидность следовых условных рефлексов. Они образуются при регулярном повторении безусловного раздражителя. Например, кормление животного через каждые 30 мин. После большого количества сочетаний (пища + определенный интервал времени) каждый раз после окончания данного интервала времени появляется рефлекторная реакция, которая ранее возникала при действии безусловного раздражителя.
Условные рефлексы на время могут быть выработаны на различные временные интервалы — от нескольких секунд до нескольких часов и даже суток. Видимо, ориентиром в отсчете времени (наличными раздражениями) могут служить различные периодические процессы, происходящие в организме (мозговых структурах — генераторы ритмов или пейсмекер, кроме того, частота сердечных сокращений, дыхательный ритм, двигательная и секреторная периодика пищеварительной системы и т. п.).
Явление отсчета времени организмом часто называют «биологическими часами». Живой организм имеет целый набор биологических часов (ритмов) с различными периодами. Короткие периоды колебаний, возникающие на клеточном уровне, трансформируются в более длительные ритмы отдельных органов и систем организма.
В зависимости от структуры условного сигнала условные рефлексы делят на простые и сложные. Иначе говоря, условными сигналами могут быть одиночные и комплексные раздражители. Среди комплексных условных рефлексов прежде всего выделяют рефлексы на одновременные и последовательные комплексы раздражителей, а также на цепь раздражителей. Так, если сигналом становится комбинация из одновременно применяемых нескольких раздражителей, то образуется условный рефлекс на одновременный комплекс раздражителей.
Например, звонок в одной экспериментальной камере является сигналом оборонительного рефлекса, а в другой камере — сигналом пищевой реакции. Очевидно, что условным сигналом в этом эксперименте служит не просто звонок сам по себе, а комплекс раздражителей, состоящий из звонка и всей обстановки опыта. Комбинация из раздражителей, которые начинают действовать друг за другом, приводит к образованию ассоциации на последовательный комплекс стимулов.
Если в комплексе сигнальных раздражителей между окончанием действия предыдущего сигнала и началом действия последующего имеется интервал времени, а подкрепление совпадает с действием только последнего компонента, то образуется условный рефлекс на цепь раздражителей.
Условные рефлексы высшего порядка. Условные рефлексы могут образовываться не только при сочетании условного сигнала с безусловным, но и при сочетании индифферентного раздражителя с условным сигналом. Например, если у собаки выработан условный слюноотделительный рефлекс на звонок, то, сочетая какой-либо индифферентный раздражитель с условным звуковым сигналом, образуется условный рефлекс II порядка, т. е. в этом случае вторичный раздражитель будет также вызывать слюноотделение, хотя ни сам по себе, ни в сочетании с первичным условным сигналом он не подкреплялся. На основе пищевой и оборонительной условной реакции у собак удается выработать условные рефлексы II и IV порядков, но при повышенном мотивационном возбуждении.
Такие «порядковые» условные рефлексы образуются на основе прочного условного рефлекса и называются рефлексами высшего порядка. Условные рефлексы высшего порядка образуются тем легче, чем более возбудима нервная система, а также чем сильнее безусловный рефлекс, на основе которого выработан рефлекс I порядка. У нервнобольных детей с патологически повышенной возбудимостью легко вырабатываются условные рефлексы V и VI порядков, в то время как у здоровых детей — только II порядка.
Условные рефлексы на цепь раздражителей следует отличать от условных цепных рефлексов, которые представляют собой специальный вид объединения двух и более условных рефлексов (синтез рефлексов). Если несколько двигательных рефлексов, выработанных на разные условные сигналы, многократно осуществляются в определенной последовательности, то сигнальный раздражитель первого в этом ряду рефлекса приобретает свойство запускать всю цепь последовательных движений, (динамический стереотип).
Сложная цепь временных связей называется динамический стереотип – этот процесс может протекать при отсутствии непосредственной связи с безусловной реакцией. Уже в лаборатории И.П. Павлова было сделано предположение, что процесс синтеза разнообразных цепных рефлексов (когда конец одного рефлекса запускает следующий рефлекс) составляет основу человеческих навыков (речевых, профессиональных, спортивных и т. п.). И.П. Павлову, его ученикам и последователям удалось воспроизвести и проанализировать эти сложные ассоциативные связи в лабораторных условиях в виде различных моделей научения на животных и человеке.
2
Процесс формирования классического условного рефлекса проходит три основные стадии.
1 Стадия прегенерализации, характеризуется концентрацией возбуждения в проекционных зонах коры условного и безусловного раздражителей.
2 Стадия генерализации условного рефлекса, в основе которой лежит процесс «диффузного» распространения (иррадиации) возбуждения. Условные реакции возникают на сигнальный раздражитель, а также интервал между предъявлениями условного стимула (межсигнальные реакции). В этот период различные биоэлектрические сдвиги широко распространены по коре и подкорковым структурам. В период генерализации условного рефлекса наблюдается синхронизация биоэлектрической активности коры и подкорковых образованиях.
3 Стадия специализации, когда межсигнальные реакции угасают и условный ответ возникает только на сигнальный раздражитель. Этот процесс обеспечивает дифференцировку и тонкое различение стимулов. В процессе специализации сфера распространения биопотенциалов значительно сужена и возрастает условно-рефлекторный ответ.
Согласно И.П. Павлову (1903), временная связь образуется между корковым центром безусловного рефлекса и корковым центром анализатора, на рецепторы которого действует условный раздражитель, т.е. связь замыкается в коре большого мозга.
В основе замыкания временной связи лежит процесс доминантного взаимодействия между возбужденными центрами. Импульсы, вызываемые условным (индифферентным) сигналом с любого участка кожи и других органов чувств (глаз, ухо), поступают в кору большого мозга и обеспечивают там образование очага возбуждения. Если после индифферентного сигнала подается пищевое подкрепление (подкормка), то возникает более мощный второй очаг возбуждения в коре больших полушарий, к которому направляется ранее возникшее и иррадиирующее по коре возбуждение.
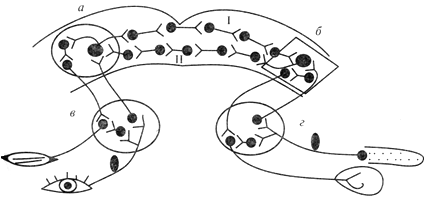
Схема дуги условного рефлекса с двусторонней связью (по Э.А. Асратяну): а – кортикальный центр мигательного рефлекса; б – кортикальный центр пищевого рефлекса; в, г – подкорковые центры мигательного и пищевого рефлексов соответственно; I – прямая временная связь; II – обратная временная связь
Неоднократное сочетание в опытах индифферентного (условного) сигнала и безусловного раздражителя (подкрепление) облегчает прохождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса. Это явления получило название синаптическое облегчение (или эффект «проторения пути»). Благодаря синаптическому облегчению формируется устойчивый очаг повышенной возбудимости нервных центров, который называется доминанта.
3
Доминанта - устойчивый очаг повышенной возбудимости нервных центров, при котором возбуждения, приходящие в центр, служат усилению возбуждения в очаге, тогда как в остальной части нервной системы наблюдаются явления торможения.
Доминанта – стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе функции других нервных центров. Явление доминанты открыл А.А. Ухтомский (1923) в опытах с раздражением двигательных зон коры большого мозга наблюдая сгибания конечности животного. Выяснилось, если раздражать корковую двигательную зону на фоне избыточного повышения возбудимости другого нервного центра, сгибания конечности может не произойти. Вместо сгибания конечности раздражение двигательной зоны может вызывать реакцию тех эффекторов, деятельность которых контролируется господствующим, т.е. доминирующим в данный момент в ЦНС, нервным центром.
В эксперименте доминанту можно получить многократной посылкой афферентных импульсов к определенному центру, гуморальными на него влияниями. Роль гормонов в образовании доминантного очага возбуждения демонстрирует опыт на лягушке: весной у самца раздражение любого участка кожи вызывает не защитный рефлекс, а усиление обнимательного рефлекса. В условиях натурального поведения доминантное состояние нервных центров может быть вызвано метаболитами.
Доминантный очаг возбуждения обладает рядом особых свойств, главными из которых являются: инерционность, стойкость, повышенная возбудимость, способность «притягивать» к себе иррадиирующие по ЦНС возбуждения и оказывать угнетающие влияния на центры-конкуренты и другие нервные центры.
Согласно Ухтомскому, доминантный очаг – это констелляция, представляющая собой «физиологическую систему», образующуюся в ходе текущей деятельности организма на всех этажах ЦНС, в разных ее участках, но с первичным фокусом возбуждения в одном из отделов и с переменным значением функций отдельных компонентов констелляции. Доминанта – общий принцип работы ЦНС, и она определяет освобождение организма от побочной деятельности во имя достижения наиболее важных для организма целей. Доминанта по всем данным «есть комплекс определенных симптомов во всем организме», проявляющийся и в мышечной, и в секреторной, и в сосудистой деятельности. Доминирует сложная система рефлексов. При «экстремальных» условиях ассоциативная связь образуется при одном-двух сочетаниях, в результате чего условные связи могут сохраняться длительное время, если не всю жизнь.
Значение доминантного очага возбуждения в ЦНС заключается в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов, необходимых для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии. Например, на базе доминантного состояния центра голода реализуется пищедобывательное поведение, на базе доминантного состояния центра жажды запускается поведение, направленное на поиск воды. Успешное завершение данных поведенческих актов, в конечном счете, устраняет физиологические причины доминантного состояния центров голода и жажды.
В настоящее время считают, что условный рефлекс сначала становится доминантой, а затем – условным рефлексом.
Образование временной связи в коре больших полушарий И.П. Павлов назвал замыканием новой условно-рефлекторной дуги: теперь подача только условного сигнала приводит к возбуждению коркового центра безусловного рефлекса и возбуждает его, т.е. возникает рефлекс на условный раздражитель – условный рефлекс.
В процессе выработки условного рефлекса образуется двусторонняя связь между двумя центрами – корковым концом анализатора, на рецепторы которого действует условный раздражитель, и центром безусловного рефлекса, на базе которого вырабатывается условный рефлекс. Это было показано в опытах, где были взяты два безусловных рефлекса: мигательный рефлекс, вызываемый струей воздуха у глаз, и безусловный пищевой. При их сочетании выработался условный рефлекс, причем если подавали воздушную струю, то возникал пищевой рефлекс, а при даче пищевого раздражителя отмечалось мигание.
Необходимым условием образования функциональной связи между корковыми центрами разных раздражителей является синхронизация биопотенциалов. Например, при сочетании света и болевого раздражения конечности, условный рефлекс возникал при высокой степени сходства биопотенциалов зрительной и двигательной областей коры.
Параллельно со становлением условного рефлекса идет процесс формирования другой условно-рефлекторной связи, специально изменяющей состояние нейронов, что выражается в росте их фоновой активности – частоты спайков («ассоциативный тонический ответ»). Если по каким-либо причинам условно-рефлекторное изменение состояния данного нейрона не возникает, то и вырабатываемый у него условный рефлекс не обнаруживается. Это дало основание для заключения, что ассоциативный ответ включает в себя формирование состояния, качественно специфического для каждой временной связи. Данное явление – один из ведущих механизмов формирования условно-рефлекторного поведения. Деятельность не существует отдельно от состояния, а выступает как единое целое с ним. Таким образом, существуют два механизма условно-рефлекторной деятельности: 1) настроечный, регулирующий состояние мозга и создающий определенный уровень возбудимости и работоспособности нервных центров и 2) запускающий, который инициирует ту или иную условную реакцию.
Эффекторное выражение условного раздражителя в процессе обучения угасает (в результате внутреннего торможения), в то же время в эфферентной части дуги подкрепляющего раздражителя возбудимость возрастает и условный стимул становится эффективным для запуска не свойственной ему ранее эффекторной реакции.
Так, сильный звуковой сигнал, вызывающий у крыс эпилептические припадки, может превратиться в условный сигнал пищевой реакции.
Взаимосвязь левого и правого полушарий при выработке условных рефлексов обеспечивается наличием комиссуральной системы (мозолистого тела, передней, задней, гиппокампальной и хабенулярной комиссур, межбугрового сращения), которая анатомически соединяет два полушария головного мозга. Получены результаты, свидетельствующие о переносе возбуждения из одного полушария в другое. Если рефлекс вырабатывается, например, на тактильное раздражение кожи одной стороны тела, то он легко воспроизводится при таком же раздражении симметричных участков кожи другой стороны тела, т.е. в процессе выработки рефлекса происходит «перенос» временной связи в другое полушарие. Предварительная перерезка мозолистого тела прекращала (или сильно затрудняла) «перенос» условного рефлекса.
Вместе с тем с помощью метода условных рефлексов было показано, что перерезка всех комиссур больших полушарий полностью не исключает возможности межполушарного взаимодействия при выработке временной связи. Эти результаты свидетельствуют о том, что элементарные формы взаимодействия двух полушарий могут осуществляться через четверохолмие и ретикулярную формацию ствола мозга.
На клеточном и молекулярном уровнях временная связь замкнется с помощью механизмов памяти. В начале выработки условного рефлекса связь осуществляется только с помощью механизмов кратковременной памяти - распространение возбуждения между двумя возбужденными корковыми центрами. По мере повторения действия условного и безусловного раздражителей и повторяющегося возбуждения соответствующих центров кратковременная память переходит в долговременную, т.е. происходят структурные изменения в нейронах.
Условные рефлексы, как отмечалось выше, изменчивы (вариабельны), они могут затормаживаться.
3
Имеются два вида торможения условных рефлексов, принципиально отличающихся друг от друга: врожденное и приобретенное, каждое из которых имеет собственные варианты.
Безусловное (врожденное) торможение условных рефлексов подразделяется на внешнее и запредельное торможение.
1. Внешнее торможение – проявляется в ослаблении или прекращении наличного (в данный момент) условного рефлекса при действии какого-либо постороннего раздражителя.
Например, включение звука, света во время текущего условного рефлекса вызывает появление ориентировочно-исследовательской реакции, ослабляющей или прекращающей наличную условно-рефлекторную деятельность. Эту реакцию, возникшую на изменение внешней среды (рефлекс на новизну), И.П. Павлов называл рефлексом «что такое?». Он состоит в настораживании и подготовке организма к действию на случай внезапно возникающей необходимости (нападение, бегство и др.). С повторением действия дополнительного раздражителя реакция на этот сигнал ослабевает и исчезает, поскольку организму не требуется предпринимать каких-то действий.
По степени выраженности влияния посторонних раздражителей на условно-рефлекторную деятельность выделяют два варианта торможения: гаснущий тормоз и постоянный тормоз.
Гаснущий тормоз – это посторонний сигнал, который с повторением его действия теряет свое тормозящее влияние, поскольку не имеет существенного значения для организма. На человека действуют множество различных сигналов, на которые сначала он обращает внимание, а затем перестает их замечать.
Постоянный тормоз – это такой дополнительный раздражитель, который с повторением не теряет своего тормозящего действия. Сюда относят раздражения от переполненных внутренних органов (от мочевого пузыря, кишечника и др.), болевые раздражители. Эти раздражители имеют существенное значение для человека и требуют от него принятия решительных мер к их устранению, поэтому условно-рефлекторная деятельность затормаживается.
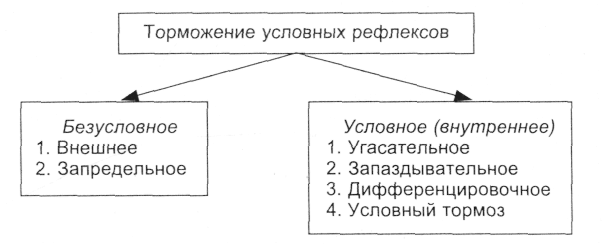
Рис. Виды торможения условных рефлексов
Механизм внешнего торможения. Согласно теории И. П. Павлова, посторонний сигнал сопровождается появлением в коре большого мозга нового очага возбуждения, который при средней силе раздражителя оказывает угнетающее влияние на текущую условно-рефлекторную деятельность по механизму доминанты. Внешнее торможение является, безусловно-рефлекторным. Поскольку в этих случаях возбуждение клеток ориентировочно-исследовательского рефлекса, возникающего от постороннего раздражителя, находится вне дуги наличного условного рефлекса, это торможение назвали внешним. Более сильный или более важный в биологическом или социальном отношении раздражитель подавляет (ослабляет или устраняет) другую реакцию. Внешнее торможение способствует экстренному приспособлению организма к меняющимся условиям внешней и внутренней среды и дает возможность при необходимости переключиться на другую деятельность в соответствии с ситуацией.
2. Запредельное торможение возникает при длительном нервном возбуждении организма, при действии чрезвычайно сильного условного сигнала или нескольких несильных. Между силой условного раздражителя и величиной ответной реакции имеется определенное соответствие – «закон силы»: чем сильнее условный сигнал, тем сильнее условно-рефлекторная реакция. Однако закон силы сохраняется до определенной величины, выше которой эффект начинает уменьшаться, несмотря на увеличение силы условного сигнала: при достаточной силе условного сигнала эффект его действия может полностью исчезнуть. Эти факты позволили И. П. Павлову выдвинуть положение о том, что корковые клетки имеют предел работоспособности. Многие исследователи относят запредельное торможение по механизму к пессимальному торможению. Поскольку появление указанного торможения не требует специальной выработки, оно, как и внешние торможения, является безусловно-рефлекторным и выполняет охранительную роль.
Условное (приобретенное, внутреннее) торможение условных рефлексов – это активный нервный процесс, который требует своей выработки, как и сам рефлекс. Поэтому его и называют условно-рефлекторным торможением: оно является приобретенным, индивидуальным.
Согласно теории И. П. Павлова, оно локализовано в пределах («внутри») нервного центра данного условного рефлекса. Различают следующие виды условного торможения: угасательное, запаздывательное, дифференцировочное и условный тормоз.
1. Угасательное торможение возникает при повторном применении условного сигнала и неподкреплении его. При этом вначале условный рефлекс ослабевает, а затем полностью исчезает, через некоторое время он может восстановиться. Скорость угасания зависит от интенсивности условного сигнала и биологической значимости подкрепления: чем они значительнее, тем труднее совершается угасание условного рефлекса. Этот процесс связан с забыванием полученной ранее информации, если она длительно не повторяется. Если во время проявления условного угасательного рефлекса подействовать посторонним сигналом, возникает ориентировочно-исследовательский рефлекс, который ослабляет угасательное торможение и восстанавливает угасший ранее рефлекс (явление растормаживания). Это показывает, что выработка угасательного торможения связана с активным угасанием условного рефлекса. Угасший условный рефлекс быстро восстанавливается при его подкреплении.
2. Запаздывательное торможение возникает при отставлении подкрепления на 1 – 3 мин относительно начала действия условного сигнала. Постепенно появление условной реакции сдвигается к моменту подкрепления. Более длительное отставление подкрепления в опытах на собаках не удается. Выработка запаздывательного условного рефлекса наиболее трудна. Этому торможению также присуще явление растормаживания.
3. Дифференцировочное торможение вырабатывается при дополнительном включении раздражителя, близкого к условному, и неподкреплении его. Например, если у собаки тон 500 Гц подкреплять пищей, а тон 1000 Гц не подкреплять и их чередовать в течение каждого опыта, то через некоторое время животное начинает различать оба сигнала: на тон 500 Гц будет возникать условный рефлекс в виде движения к кормушке, поедания корма, слюноотделения, а на тон 1000 Гц животное будет отворачиваться от кормушки с пищей, слюноотделения не будет. Чем меньше различия между сигналами, тем труднее идет выработка дифференцировочного торможения. Удается выработать у животных различение частот метронома – 100 и 104 уд./мин, тонов 1000 и 995 Гц, геометрических фигур, а также различение области раздражения различных участков кожи и т.д. Условное дифференцировочное торможение при действии посторонних сигналов средней силы ослабевает и сопровождается явлением растормаживания, т.е. это такой же активный процесс, как и при других видах условного торможения.
4. Условный тормоз возникает при добавлении к условному сигналу другого раздражителя и неподкреплении этой комбинации. Так, если выработать условный слюноотделительный рефлекс на свет, затем к условному сигналу «свет» подключить дополнительный раздражитель, например «звонок», и не подкреплять эту комбинацию, то на нее постепенно условный рефлекс угасает. Сигнал «свет» необходимо продолжать подкреплять пищей или вливанием в рот слабого раствора кислоты. После этого присоединение сигнала «звонок» к любому условному рефлексу ослабляет его, т.е. «звонок» стал условным тормозом для любого условного рефлекса. Этот вид торможения также растормаживается, если подключить другой раздражитель.
Значение всех видов условного (внутреннего) торможения условных рефлексов заключается в устранении ненужной в данное время деятельности – тонком приспособлении организма к окружающей среде.
Механизм условного (внутреннего) торможения условных рефлексов изучен недостаточно. При развитии условного торможения в этот процесс широко вовлекаются различные мозговые структуры. Ученые полагают, что в основе угасания лежит механизм усиления тормозящих влияний на ранее возбуждавшиеся клеточные элементы системы условного рефлекса. В модельных ситуациях обнаружено, что при угасательном торможении амплитуда возбуждающего постсинаптического потенциала постепенно снижалась
На клеточном уровне это, по-видимому, осуществляется с помощью механизмов памяти. Торможение условных рефлексов можно объяснить тем, что при неподкреплении условного раздражителя исключаются доминантные взаимоотношения двух очагов возбуждения, остается только возбуждение от условного сигнала. Поэтому проторенный ранее путь между доминантными очагами возбуждения постепенно устраняется, так как исчезает синаптическое облегчение, сформированное ранее.