

1.Строение плазматической мембраны, роль ионных каналов, молекул - переносчиков, насосов, рецепторов. Виды транспорта веществ через мембрану клетки. Плазматическая мембрана- эластическая структура, толщиной от 7 до 11 нм. Она состоит, в основном, из белков и липидов (40-90% фосфолипиды- фосфотидилхолин, фосфотидилэтаноламин, фосфотидилсерин, сфингомиелин, фосфотидилинозит. Так же, важным компонентом мембран являются гликолипиды- цереброзиды, сульфатиды, ганглиозиды и холестерин). Основной структурой мембраны клетки является двойной слой фосфолипидных молекул, взаимодействующих с белковыми молекулами. Белки мембран клеток представлены, в основном, гликопротеинами. Различают интегральные белки, проникающие через всю толщу мембраны и периферические белки, прикреплённые только к поверхности мембраны, в основном, к внутренней её части. Периферические белки почти все функционируют как энзимы, а интегральные белки обеспечивают селективный обмен ионов через каналы мембран между экстрацелюлярной и интрацелюлярной жидкостью. Ионный канал (ИК) - это интегральный белок, образующий в мембране пору для обмена клетки с окружающей средой ионами K+, Na+, H+, Ca2+, Cl-, а также водой, и способный изменять свою проницаемость. Транспорт: А) пассивный А1 - простая диффузия - основана на разности концентраций веществ внутри клетке и в меж клеточном пространстве. Возникает поток частиц в сторону с меньшей концентрацией. Поток прекращается после выравнивание концентраций. Через кл. мембр. проходят и гидрофильные и гидрофобные вещества. Первые проходят растворённые в воде, а вторые диффундируют благодаря растворению в липидах мембраны. А1.1- осмос- движение молекул воды через мембрану из области большей концентрации в обл. с меньшей концентрацией растворённого в ней вещества. А1.2. –Диффузия ионов – происходит через ИК (натриевые, калиевые, кальциевые, хлорные, натрий- кальциевые). Каналы могут быть в закрытом, открытом и инактивированном состоянии, Переход из 1 состояние в другое осущ. изменением электрической разности потенциалов или взаимодействием рец. с активными веществами. А2 – Облегчённая диффузия - При облегченной диффузии вещества переносятся через мембрану также по градиенту концентрации, но с помощью специальных трансмембранных белков-переносчиков (транслоказ). Белок-переносчик имеет центр связывания, комплементарный переносимому веществу, поэтому для облегченной диффузии, в отличие от простой, характерна высокая избирательность: для каждого вещества или группы сходных веществ имеется свой переносчик. Б) активный- Типы активного транспорта. Первичный активный транспорт – осущ транспортными АТФа- зами – ионными насосами (интегральный белок+ Ca –АТФазы) за счёт энергии гидролиза АТФ.Вторично активный тр.- перенос веществ через мембр. против градиента конц. за счёт энергии градиента конц. Другого вещества, создаваемого в процессе активного транспорта (напр. В кардиомиоцитах с функционированием Na, K – АТФазы связан транспорт Ca)
2.Свойства живых и возбудимых систем: раздражимость, возбудимость, проводимость, лабильность, их количественные показатели. Сравнительная оценка возбудимости тканей. Раздражимость — общее свойство любой живой ткани или клетки реагировать на внешнее воздействие изменением своих физико- химических и физиологических свойств. Проявляется в изменении текущих значений физиологических параметров, величина которых превышает их сдвиги при покое. Присуща всем биологическим системам.
Возбудимость- способность организма, органа, ткани или клетки отвечать на раздражение активной специфической реакцией – возбуждением (генерация нервного импульса, сокращение, секреция). Возбудимость различных тканей не одинакова. Её величину оценивают по порогу раздражения – минимальная сила раздражителя, способного вызвать возбуждение.
Критическая точка возбуждения у нервной и мышечной клетки находится на уровне – 50 мВ, но потенциал покоя у этих клеток разный( у нервной – 70 мВ, у мышечной -90мВ), из за этого пороговое возбуждение у нервной ткани меньше( у нервной 20 мВ, у мышечной 40 мВ), следовательно, нервная ткань является более возбудимой.
Лабильность (функциональная подвижность) - способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений. Это свойство характеризует скорость возникновения возбуждения. Показатель лабильности: максимальное количество волн возбуждения в данной ткани: нервные волокна - 500-1000 импульсов в секунду, мышечная ткань - 200-250 импульсов в секунду, синапс - 100-125 импульсов в секунду. Лабильность зависит от уровня обменных процессов в ткани, возбудимости, рефрактерности.
Проводимость - способность ткани проводить возбуждение по всей своей длине. Показатель проводимости - скорость проведения возбуждения. Скорость проведения возбуждения по скелетной ткани - 6-13 м/с, по нервной ткани до 120 м/с. Проводимость зависит от интенсивности обменных процессов, от возбудимости (прямо пропорционально).
3. Потенциал покоя и потенциал действия: их происхождение, фазы потенциала действия.
Имеет калиевую природу. В покое в мембране кл. калиевые каналы - открыты, натриевые каналы- закрыты, т.к. внутри кл. калия больше, он по градиенту концентрации выходит наружу, унося с собой положительный заряд. Вслед за уходящим калием, к внутренней поверхности мембраны устремляются ионы хлора. В результате изнутри мембрана заряжается “-”, а снаружи “+”. Натрий калиевые АТФазы возвращают калий обратно в кл., но в обмен на 2 иона калия, они выносят наружу 3 иона натрия, т.е. утечка “+” заряда из кл. продолжается и на мембране формируется стойкая разность потенциалов. Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны.
Имеет натриевую природу. Когда на кл. действует раздражитель, постепенно начинают открываться натриевые каналы. Заходящие в кл ионы натрия понижают разность потенциалов на мембране. У каждой возбудимой кл. есть критический уровень деполяризации. Когда разность потенциалов достигает этого уровня, все натриевые каналы в мембране- открываются. Натрий лавинообразно устремляется внутрь кл. и происходит перезарядка мембраны.
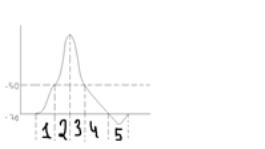
Фаза медленной деполяризации,
фаза быстрой деполяризации,
фаза реполяризации,
следовая реполяризация.
следовая гиперполяризация
4) Фазные изменения возбудимости в их сопоставлении с компонентами потенциала действия
Импульсное возбуждение сопровождается фазными изменениями возбудимости:
фаза кратковременного повышения возбудимости (рефрактерный период делится на абсолютный и относительный);
фаза супернормальной возбудимости;
фаза субнормальной возбудимости.
Фаза кратковременного повышения возбудимости соответствует местному компоненту потенциала действия, т. к. происходит частичное повышение проницаемости для Na+, что приводит к критическому уровню деполяризации (Ек). Уменьшается порог раздражения и повышается возбудимость, местное возбуждение всегда сопровождается повышением возбудимости.
ткань приходит в состояние готовности ответить на раздражение.
Рефрактерный период делится на:
абсолютный - соответствует фазе деполяризации (максимально открыты Nа-каналы и дальнейшее увеличение проницаемости для Nа невозможна). Значение: совершается основная работа;
относительный - возбудимость восстанавливается до исходной величины (заряд клеточной мембраны возвращается к исходной величине). Закрываются Nа-каналы, поэтому можно получить дополнительную реакцию при действии сверхпорогового раздражителя. Значение: в этот момент возможно получить ответ на сильные биологически важные раздражители.
Фаза супернормальной возбудимости (экзальтации) соответствует отрицательному следовому потенциалу, Nа-каналы еще не все закрыты, поэтому достаточно более слабого раздражителя для получения ответной реакции.
Фаза субнормальной возбудимости. Разность потенциала увеличивается, увеличивается состояние до Ек. Порог раздражения увеличивается, возбудимость снижается.
5 Классификация раздражителей. Законы раздражения: закон силы для одиночных клеток, закон силы для группы клеток, закон соотношения силы и времени, закон градиента.
Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.
Раздражители – это факторы внешней или внутренней среды, действующие на возбудимые структуры.
Различают две группы раздражителей:
1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах);
2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т. п.), физико-химические (осмотические – кристаллик хлорида натрия).
Классификация раздражителей по биологическому принципу:
1) адекватные, которые при минимальных энергетических затратах вызывают возбуждение ткани в естественных условиях существования организма;
2) неадекватные, которые вызывают в тканях возбуждение при достаточной силе и продолжительном воздействии.
По силе:
Пороговые, Надпороговые, Подпороговые, Сверхпороговые
Законы раздражения возбудимых тканей
Законы устанавливают зависимость ответной реакции ткани от параметров раздражителя. Эта зависимость характерна для высоко организованных тканей. Существуют три закона раздражения возбудимых тканей:
1) закон силы раздражения;
2) закон длительности раздражения;
3) закон градиента раздражения.
Закон силы раздражения устанавливает зависимость ответной реакции от силы раздражителя. Эта зависимость неодинакова для отдельных клеток и для целой ткани. Для одиночных клеток зависимость называется «все или ничего». Характер ответной реакции зависит от достаточной пороговой величины раздражителя. При воздействии подпороговой величиной раздражения ответной реакции возникать не будет (ничего). При достижении раздражения пороговой величины возникает ответная реакция, она будет одинакова при действии пороговой и любой сверхпороговой величины раздражителя (часть закона – все).
Для совокупности клеток (для ткани) эта зависимость иная, ответная реакция ткани прямо пропорциональна до определенного предела силе наносимого раздражения. Увеличение ответной реакции связано с тем, что увеличивается количество структур, вовлекающихся в ответную реакцию.
Закон длительности раздражений. Ответная реакция ткани зависит от длительности раздражения, но осуществляется в определенных пределах и носит прямо пропорциональный характер. Существует зависимость между силой раздражения и временем его действия. Эта зависимость выражается в виде кривой силы и времени. Эта кривая называется кривой Гоорвега—Вейса—Лапика. Кривая показывает, что каким бы сильным ни был бы раздражитель, он должен действовать определенный период времени. Если временной отрезок маленький, то ответная реакция не возникает. Если раздражитель слабый, то бы как длительно он ни действовал, ответная реакция не возникает. Сила раздражителя постепенно увеличивается, и в определенный момент возникает ответная реакция ткани. Эта сила достигает пороговой величины и называется реобазой (минимальной силой раздражения, которая вызывает первичную ответную реакцию). Время, в течение которого действует ток, равный реобазе, называется полезным временем.
Закон градиента раздражения. Градиент – это крутизна нарастания раздражения. Ответная реакция ткани зависит до определенного предела от градиента раздражения. При сильном раздражителе примерно на третий раз нанесения раздражения ответная реакция возникает быстрее, так как она имеет более сильный градиент. Если постепенно увеличивать порог раздражения, то в ткани возникает явление аккомодации. Аккомодация – это приспособление ткани к медленно нарастающему по силе раздражителю. Это явление связано с быстрым развитием инактивации Na-каналов. Постепенно происходит увеличение порога раздражения, и раздражитель всегда остается подпороговым, т. е. порог раздражения увеличивается.
Законы раздражения возбудимых тканей объясняют зависимость ответной реакции от параметров раздражителя и обеспечивают адаптацию организмов к факторам внешней и внутренней среды.
7вопрос!!!
Отличия ПКП от ПД:
ПКП в 10 раз дольше ПД. ПКП возникает на постсинаптической мембране. ПКП обладает большей амплитудой. Величина ПКП зависит от числа молекул ацетилхолина, связанных с рецепторами постсинаптической мембраны, т.е. в отличие от потенциала действия ПКП градуален.
6Классификация нервных волокон, особенности проведения возбуждения по миелиновым и безмиелиновым волокнам. Законы проведения возбуждения по нервным волокнам.
Главная функция нервных волокон – проведение нервных импульсов. Отростки нервных клеток проводят сами нервные импульсы, а глиальные клетки способствуют этому проведению. По особенностям строения и функциям нервные волокна подразделяются на два вида: безмиелиновые и миелиновые.
Безмиелиновые нервные волокна не имеют миелиновой оболочки. Их диаметр 5–7 мкм, скорость проведения импульса 1–2 м/с. Миелиновые волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и оксоплазму. Миелиновая оболочка состоит на 80 % из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье). Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. При диаметре 12–20 мкм скорость проведения возбуждения составляет 70—120 м/с.
В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа: А, В, С.
Наибольшей скорость проведения возбуждения обладают волокна типа А, скорость проведения возбуждения которых достигает 120 м/с, В имеет скорость от 3 до 14 м/с, С – от 0,5 до 2 м/с.
Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну.
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.