

Картирование с использованием транслокаций и делеций
Используется гибрид клетки и хромосома с хромосомными перестановками.
Транслокация – хромосомная перестройка, в ходе которой хромосомы обмениваются участками, причем негомологичными. Было установлено, что три гена локализованы в Х-хромосоме, но в каком месте – неизвестно.
Э ти
клетки гибридизовались клетками
мыши, и далее отобрали выделенный клон,
в котором соединились обе перестроенные
хромосомы.
ти
клетки гибридизовались клетками
мыши, и далее отобрали выделенный клон,
в котором соединились обе перестроенные
хромосомы.
Клоны ► |
Х/14 14/Х |
Х/14 |
14/Х |
ГГФРТ ФКГ Г-6ФД |
+ + + |
- - - |
+ + + |
Клоны ► |
|
|||
ГГФРТ ФКГ Г-6ФД |
+ + + |
+ - + |
- - + |
- - - |

Картирование генов у человека: метод днк-зондов.
ДНК-зонд – «капля» целого гена или его части, помеченная радиоактивным или флуоресцентным методом. Наиболее часто получают методом обратной транскрипции. Была выделена из эритроцитов и-РНК, кодирующая α,β-цепи гемоглобина, и на базе этой РНК получили меченые зонды. Далее взяли культуру клеток человека и приготовили из нее хромосомные препараты. На одном из этапов клетки были обработаны, с целью вызвать неполную денатурацию, и в это же время было добавлено большое количество зондов. Так как зондов много, то они могут заменять участки ДНК, к которым они комплиментарны, далее цепи ренатуируют. Когда хромосомные препараты были приготовлены и окрашены, то на том участке, где к хромосоме присоединился зонд, будет наблюдаться флуоресценция. Если хромосома хорошо дифференцируема, то можно определить, какая это хромосома и на каком участке произошло присоединение.
Митотический цикл клетки. Характеристика его периодов.

54. Митоз и его биологическое значение. Проблемы клеточной пролиферации в медицине.
Митоз состоит из 4 фаз - профаза, метафаза, анафаза, телофаза. Профаза. Первыми процессами профазы является процесс компактизации хромосом. Чтобы хромосомы нормально разошлись, они должны резко сократиться в размерах. Например, первая хромосома имеет длину 2 см, в конце профазы 10 микрон. Исчезает ядрышко, в котором происходит биосинтез рРНК. Наблюдается исчезновение ядерной оболочки. В этот момент центриоли клеточного центра начинают движение к разным полюсам и между ними натягивается межполюсной ахроматиновый аппарат. Некоторые ученые выделяют прометафазу (промежуточная стадия). Она характеризуется тем, что от центриолей к хромосомам начинают двигаться хромосомные нити и как только нить касается кинетохора - она начинает толкать нить к противоположному полюсу, до того как с противоположного полюса не образуется другая нить, которая также прикрепляется к кинетохорам и толкает их. Это вот толкание - прометафаза. Метафаза. Стадия, когда все хромосомы выстраиваются на экваторе клетки и каждой хромосоме от полюсов прикрепляется нить ахроматинового аппарата. Существую ещё межполюсные нити. Если смотреть в микроскоп, то мы увидим конфигурацию Материнская звезда. Свободные концы хромосом наружу. Метафаза заканчивается тем, что центромерная область хромосомы расщепляется и половины хромосом - хроматиды начинают движение к разным полюсам, при этом движение хромосом определяет особый белок - динеин, который располагается в области кинетохора хромосом. Происходит укорочение хромосомных нитей за счет деполимеризации волокна в области центриолей, то есть тубулиновые вставки начинают разваливаться, удаляются и волокно становится короче, то есть, доказаны 2 фактора, определяющие движение хромосомы: Моторный белок динеин и разборка тубулинового волокна. Однако тонкость этого механизма непонятна. Анафаза. Начинается с расхождения хроматид. И до того момента, когда однохроматидная хромосома достигнет противоположного плюса. Длится 10-15 секунд. Телофаза. Начинается с момента полного отхождения хромосом к полюсам и заканчивается началом формирования дочерних ядер и образованием перегородки между этими ядрами. Это называется цитокинез или цитомия. Когда хромосомы достигнут противоположных полюсов наблюдается явление деспирализации хромосом и образование структур, характерных для интерфазой клетки, например ядрышка, а также энодплазматической сети, так как она исчезает. Цитокинез различен у растительных и животных клеток. У животных происходит впячивание вовнутрь клеточной стенки в виде гантели. У растения же клеточная стенка образуется в центре и движется к границам клетки. Таким образом, благодаря митозу образуется 2 дочерних клетки с идентичным генетическим материалом
Значение митоза: обеспечивает равномерное распределение хромосом между дочерними клетками.
Мейоз и его биологическое значение
Мейоз. Впервые в 1884 Ван Бенеден открыл и описал мейоз, но еще до него немецкий ученый Август Вейсман доказал, что в половых клетках должно происходить уменьшение числа хромосом вдвое, то есть редукция хромосом. В мейозе различают редукционное и эквационное уравнительное деление. Редукционное деление имеет 4 стадии - профазу, метафазу, анафазу, телофазу. К моменту Профазы I каждая хромосома двухроматидна и имеет 4с2n. Профаза делится на 5 фаз: 1. Леитотена 2. Зиготена 3. Пахинема 4. Диплотена 5. Диакинез. Лептонема. Стадия клубка нитей. Хромосомы спирализуются, образуя клубок, который формируется около одной из ядерных стенок. При этом свободные концы ориентированы в направлении клеточного центра. Зигонема. Происходит дальнейшее уплотнение и укорочение хромосом. При этом гомологические хромосомы, то есть хромосомы, которые несут одинаковые ген, одна из которых пришла с отцовской, а другая с материнской, объединяются - происходит конъюгация. Образуется бивалент. Иногда её называют тетрадой, так как здесь 4 хроматиды. В этот момент центриоли начинают движение к разным полюсам - формирование межполюсных нитей. Ядерная оболочка начинает исчезать. Пахинема. Здесь происходит процесс отталкивания хромосом и формирование хиазм - переплетение между хроматидами гомологичных хромосом. Диплонема. Хромосомы начинают ещё дальше отходить друг от друга, а в местах хиазм происходит обмен участками. Хромосомы становятся гибридными - рекомбинация генетического материала - кроссинговер. Диакинез. В диакинезе хромосомы ещё удерживаются друг относительно друга своими концевыми участками. К центромерам хромосом подходят центромерные нити, которые идут от клеточного центра. В отличие от митоза к хромосоме при мейозе прикрепляется 1 нить. Метафаза I. Хромосомы располагаются на экваторе. К каждой двухроматидной хромосоме прикрепляется нить веретена. Анафаза I. Целые хромосомы начинают движение к разным полюсам. Телофаза I - формирование двух дочерних ядер - 1n хромосом, двухроматидных хромосом. Далее идет интеркинез, в котором биосинтеза ДНК не происходит. Эквационное деление. Профаза I. Хромосомы спирализуются, укорачиваются, ядерная оболочка растворяется. Центриоли расходятся к полюсам, натягивается аппарат деления. Метафаза I. Двухроматидные хромосома но экваторе. К центромерам подходят нити.
Анафаза. Хроматида движется к полюсам. Телофаза. Здесь формируется 2 ядра. Происходит цитокинез. 56. СПЕРМАТОГЕНЕЗ
Сперматогенез - процесс развития мужских половых клеток, заканчивающийся формированием сперматозоидов. Протекает внутри извитых семенных канальцев, составляющих более 90% объёма яичка взрослого половозрелого мужчины.
На внутренней стенке канальцев располагаются клетки 2 типов - сперматогонии (самые ранние, первые клетки сперматогенеза, из которых в результате последовательных клеточных делений через ряд стадий постепенно образуются зрелые сперматозоиды) и питающие клетки Сертоли.
Сперматогенез начинается одновременно с деятельностью яичка под влиянием половых гормонов в период полового созревания подростка и далее протекает непрерывно (у большинства мужчин практически до конца жизни), имеет чёткий ритм и равномерную интенсивность.
Время, необходимое для превращения сперматогония в спермий, занимает у человека около 74-75 суток. При этом сперматогонии, которые встречаются в яичках мальчиков ещё до наступления периода полового созревания, бывают двух типов: А и В, или тёмные и светлые; часть из них сохраняется в качестве запасных, а другие начинают расти и делиться.
Сперматогонии, содержащие удвоенный набор хромосом, делятся путём митоза, приводя к возникновению последующих клеток - сперматоцитов 1-го порядка. Далее в результате двух последовательных делений (мейотические деления) образуются сперматоциты 2-го порядка, а затем сперматиды (клетки сперматогенеза, непосредственно предшествующие сперматозоиду).
При этих делениях происходит уменьшение (редукция) числа хромосом вдвое. Сперматиды не делятся, вступают в заключительный период сперматогенеза (период формирования спермиев) и после длительной фазы дифференцировки превращаются в сперматозоиды.
Происходит это путём постепенного вытяжения клетки, изменения, удлинения её формы, в результате чего клеточное ядро сперматида образует головку сперматозоида, а оболочка и цитоплазма - шейку и хвост. В последней фазе развития головки сперматозоидов тесно примыкают к клеткам Сертоли, получая от них питание до полного созревания.
После этого сперматозоиды, уже зрелые, попадают в просвет канальца яичка и далее в придаток, где происходит их накопление и выведение из организма во время семяизвержения.
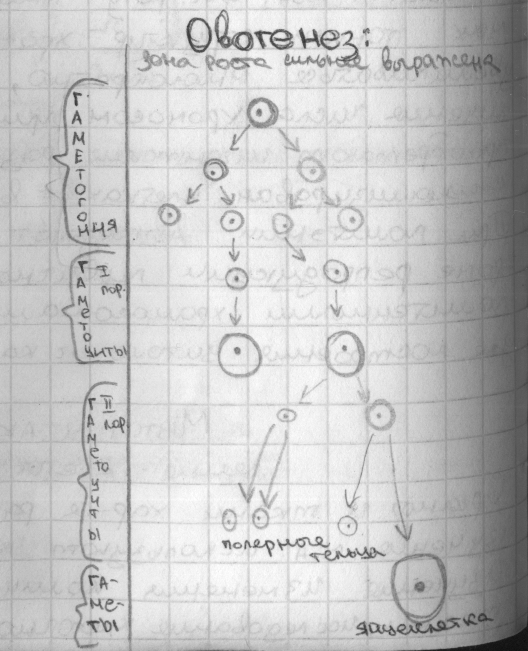
57. Овогенез. Цитологические и цитогенетические характеристики.
Овогенез — это процесс образования женских половых гамет, идет по той же схеме, что и сперматогенез, но с некоторыми существенными отличиями.
В
результате неравномерного распределения
цитоплазмы как при первом, так и при
втором делениях мейоза только в одной
клетке оказывается большой запас
питательных веществ, необходимых для
развития будущего зародыша. Следовательно,
образуется только одна зрелая яйцеклетка
с гаплоидным набором хромосом (n) и три
маленькие клеточки, которые впоследствии
исчезают. При овогенезе наряду с мейозом
происходит так называемое созревание
яйцеклетки, во время которого значительно
увеличивается ее объем
58. Взаимодействие аллелей в детерминации признаков: доминирование, промежуточное проявление, рецессивность, кодоминирование. Множественный аллелизм.


59. Взаимодействие неаллельных генов. Комплементарность.
Комплементарность — вид взаимодействия неаллельных генов, при котором признак формируется в результате суммарного сочетания продуктов их доминантных аллелей. Имеет место при наследовании ореховидной формы гребня у кур, синей окраски баклажанов, зеленого оперения у волнистых попугайчиков и пр.
Ореховидная форма гребня у кур обусловливается взаимодействием двух доминантных аллелей комплементарных генов А и В(А_В_). Сочетание одного из этих генов в доминантном, а другого в рецессивном состоянии вызывает формирование либо розовидного (А_bb), либо гороховидного гребня (ааВ_). У особей с генотипом aabb — листовидный гребень.
P |
♀АaBb ореховидный |
× |
♂AаBb ореховидный |
Типы гамет |
|
|
AB Ab aB ab |
|
♂ |
AB |
Ab |
aB |
ab |
♀ |
|
||||
AB |
AABB ореховидный |
AABb ореховидный |
AaBB ореховидный |
AaBb ореховидный |
|
Ab |
AABb ореховидный |
AАbb розовидный |
AaBb ореховидный |
Aabb розовидный |
|
aB |
AaBB ореховидный |
AaBb ореховидный |
aaBB гороховидный |
aaBb гороховидный |
|
ab |
AaBb ореховидный |
Aabb розовидный |
aaBb гороховидный |
aabb листовидный |
|
Ореховидная форма гребня — 9/16, розовидная форма гребня — 3/16, гороховидная форма гребня — 3/16, листовидная форма гребня — 1/16. Расщепление по фенотипу 9:3:3:1.
Темно-синяя окраска плодов баклажанов формируется в результате взаимодействия продуктов двух неаллельных доминантных генов А и В. Растения, гомозиготные по любому из соответствующих рецессивных аллелей а и b или по ним обоим, имеют белые плоды.
|
♂ |
AB |
Ab |
aB |
ab |
♀ |
|
||||
AB |
AABB синяя |
AABb синяя |
AaBB синяя |
AaBb синяя |
|
Ab |
AABb синяя |
AАbb белая |
AaBb синяя |
Aabb белая |
|
aB |
AaBB синяя |
AaBb синяя |
aaBB белая |
aaBb белая |
|
ab |
AaBb синяя |
Aabb белая |
aaBb белая |
aabb белая |
|
Синяя окраска плодов у баклажанов — 9/16, белая окраска плодов у баклажанов — 7/16. Расщепление по фенотипу 9:7. При комплементарном действии генов расщепление по фенотипу может быть не только 9:3:3:1 и 9:7, как в приведенных выше примерах, но и 9:6:1 и 9:3:4.
60 Взаимодействие неаллельных генов. Эпистаз, его виды.
Эпистаз — вид взаимодействия неаллельных генов, при котором одна пара генов подавляет (не дает проявиться в фенотипе) другую пару генов.
Гены, подавляющие действие других, не аллельных им генов, называются Эпистатичными, А подавляемые — Гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию. При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном.
Эпистаз может быть доминантным и рецессивным.
При доминантном эпистазе, когда доминантный аллель одного гена (А) препятствует проявлению аллелей другого гена (B или b), расщепление в потомстве зависит от их фенотипического значения и может выражаться соотношениями 12:3:1 или 13:3. При рецессивном эпистазе ген, определяющий какой-то признак (В), не проявляется у гомозигот по рецессивному аллелю другого гена (аа). Расщепление а потомстве двух гетерозигот по таким генам будет соответствовать соотношению 9:3:4. Невозможность формирования признака при рециссивном эпистазе расценивают также как проявление несостоявшегося комплементароного взаимодействия, которое возникет между доминантным аллелем эпистатического гена и аллелями гена, определяющего этот признак.