

37.Сопряженные изменения рождаемости и смертности в популяциях. Принцип Олли.
Сопряженные
изменения рождаемости и смертности.Как
неоднократно отмечалось выше,
рождаемость и смертность в популяциях
изменяются под воздействием многочисленных
факторов. Рассмотрим два
возможных случая их изменения с
увеличением плотности популяции.
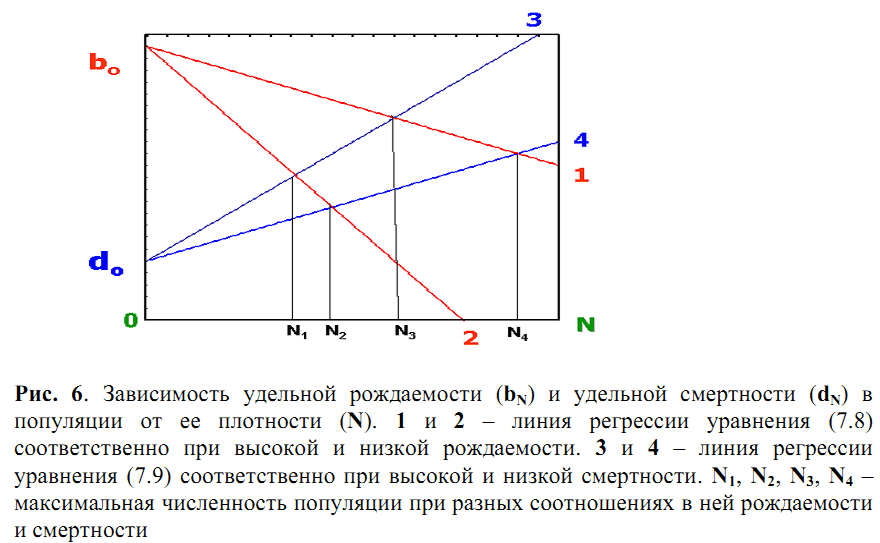
В первом случае (рис. 6) удельная скорость смертности (d) линейно возрастает, а удельная скорость рождаемости (b) линейно снижается с увеличением плотности популяции (N). Тогда связь b и N следует уравнению
BN = b0– xN, (7.8)
а связь d и N – уравнению
dn= d0+ yN, (7.9)
где bN и dN– значения удельных скоростей рождаемости и смертности при
плотности N, b d их значения при плотности, близкой к нулю, х и у – эмпирические коэффициенты. Когда bN = dN, что соответствует точке пересечения линий регрессии (7.8) и (7.9) на графике, скорость роста численности популяции (ΔN/Δt) становится равной нулю. Отсюда популяция достигает максимально возможной численности (Nmax) при данных условиях среды, или т.наз. «стационарного состояния». Легко видеть, что чем сильнее рождаемость снижается, а смертность возрастает с увеличением плотности, тем более низкими становятся значения Nmax.


![]()


38.Демографические показатели популяций, жизненные таблицы.






![]()
![]()

39.Многолетняя динамика численности популяций и методы ее оценки.
40.Межпопуляционные взаимоотношения и их классификация. Нейтрализм как форма взаимодействия.


41.Хищничество и его значение в экосистемах. Модель Лотки – Вольтерра.
Хищничество (+ : -). Для взаимодействий типа «хищник – жертва» итальянские математики А. Лотка в 1925 г. и независимо В. Вольтерра в 1926 г. разработали первую математическую модель экологического процесса, ставшую первой подобной моделью в биологической
науке. Методологические подходы, использованные для разработки модели «хищник – жертва», стали основой для моделирования других типов межпопуляционных взаимодействий.
Отношения типа «хищник – жертва» чрезвычайно распространены в природе. Они не далеко не ограничиваются хрестоматийными примерами, такими как «волки – овцы» или «кошки – мыши». В широком смысле они охватывают все межпопуляционные взаимодействия, при которых одни организмы используют в пищу другие, при этом умерщвляя их. Поэтому к ним
также относятся такие распространенные в природе связи, как «растительноядные животные – растения», например, «дафнии – одноклеточные водоросли». Хищниками являются также киты, фильтрующие планктон, и птицы, питающиеся семенами. Целый ряд отношений, обычно включаемых в категорию «паразит – хозяин», скорее относятся к типу «хищник – жертва». Так, свободноживущие перепончатокрылые насекомые из семейства наездников (Ichneumonidae)
откладывают яйца в тела личинок и куколок других видов насекомых. Выходящие из яиц личинки используют в пищу ткани личинок и куколок и к концу развития приводят своих хозяев к гибели. Подобные виды носят название паразитоидов. Поэтому сейчас в экологии часто говорят не об отношениях «хищник – жертва», а об отношениях «потребитель – ограниченный пищевой ресурс» или «жертва – эксплуататор». В этих отношениях потребителем (эксплуататором) является гетеротрофный организм, а его жертвой, или пищевым ресурсом, –
любой другой живой организм. Однако иногда потребителем может являться даже растение, например насекомоядная росянка. Иногда, в случае каннибализма, потребитель и потребляемая им пища относятся к одному виду.
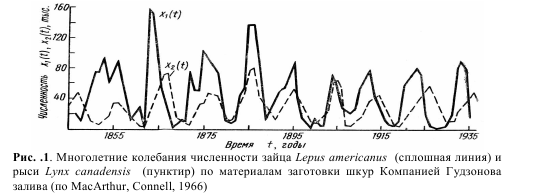
Модель Лотки – Вольтерра. Толчком к разработке этой модели стал анализ статистических данных о заготовках пушнины Компанией Гудзонова залива с конца XVIII и до середины XX века на территории современной Канады. Поскольку промысел пушнины в то время был очень интенсивным, можно считать, что количество заготовленных шкур промысловых видов
пропорционально их численности в природе. Было замечено, что численность основных объектов промысла – американского зайца-беляка Lepus americanus и охотящейся на него канадской рыси Lynx canadensis претерпевала значительные циклические колебания с периодом около 10 лет (рис.1). При этом максимумы численности хищника, как правило, наблюдались на 2–3 года позднее максимумов численности жертвы.
Рассмотрим простейший вариант модели Лотки – Вольтерры. В ней предполагается, что увеличение численности каждой из взаимодействующих популяций (хищника и жертвы) ограничивается только другой популяцией. Поэтому рост численности жертв лимитируется их потреблением хищниками, а рост численности хищников – недостаточным числом жертв. Принимается, что количество пищи для жертвы всегда достаточно, т. е. численность ее
популяции никогда не превышает емкость среды. Поэтому никакого самоограничения численности популяций жертвы или хищника по принципу отрицательной обратной связи модель не предполагает.
Обозначим численность популяции жертвы как N1, а популяции хищника – как N2. Допустим, численность популяции жертвы в отсутствии хищника растет экспоненциально, а пресс хищника тормозит этот рост. При этом смертность в популяции жертвы прямо пропорциональна частоте встреч хищника и жертвы, т.е. произведению плотностей их популяций. Тогда мгновенная скорость изменения численности жертвы (dN1/dt) может быть
выражена уравнением
dN1/dt = r1N1 – p1N1N2, (1)
где r1 – удельная скорость роста численности жертвы, p1 – константа, связывающая смертность жертвы с плотностью хищника. Очевидно, p1 > 0. Мгновенная скорость роста численности популяции хищника (dN2/dt) принимается равной рождаемости, которая, в свою очередь, зависит от интенсивности потребления хищников жертв и удельной скорости смертности:
dN2/dt = p2N1N2 - d2N2, (2)
где p2 – константа, связывающая удельную рождаемость в популяции хищника с плотностью жертв, d2 – удельная скорость смертности в популяции хищника. Решение системы этих уравнений позволяет определить условия поддержания постоянной, или равновесной численности хищника и жертвы. Если в (2) принять, что dN1/dt = 0, тогда r1N1 = p1N1N2. Отсюда популяция жертвы сохраняет постоянную численность, если численность популяции
хищника равна N2 = r1/p1. Аналогичным образом доказывается, что популяция хищника сохраняет постоянную численность, если численность популяции жертвы равна N1 = d2/p2.
Отложим на графике по оси абсцисс численность жертв (N1), а по оси ординат – численность хищников (N2). Тогда линии (изоклины), показывающие условия постоянства численности хищника и жертвы, представляют две прямые, перпендикулярные друг другу и координатным осям (рис. 2а). Точка их пересечения, соответствующая равной численности хищника и жертвы (N1 = N2), следует условию r1/p1 = d2/p2. При этом если численность жертвы N1 < d2/p2, то численность хищника (N2) всегда будет уменьшаться, а если N1 > d2/p2 – возрастать. Соответственно и численность жертвы возрастает, если численность хищника N2 < r1/p1, и
уменьшается – если N2 > r1/p1. Решение системы уравнений (1) и (2) позволяет получить зависимость числа хищников от числа жертв (рис. 2), или так называемый «фазовый
портрет системы». Она представляет собой окружность, центр которой образует пересечение изоклин хищника и жертвы. Образованный окружностью круг изоклины делят на четыре сектора. В первом секторе наблюдается возрастание численности хищника и жертвы, во втором – численность жертвы уменьшается, хищника - увеличивается, в третьем – уменьшение численности обоих видов, а в четвертом – уменьшение численности хищника и увеличение
численности жертвы.
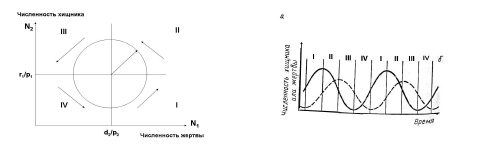
Рис. 2. Графическая модель, показывающая возникновение устойчивых колебаний
численности хищника и жертвы. а. положение изоклин, характеризующих постоянство
численности хищника и жертвы. Стрелками показаны векторы изменения численности
хищника и жертвы. б. Циклические колебания численности хищника (пунктир) и жертвы
(сплошная линия), возникающие согласно данной модели.
Радиус полученного круга (R), отражающий размах колебаний численности обоих взаимодействующих видов, легко рассчитать по теореме Пифагора:
R2 = (ΔN1)2 + (ΔN2)2 (3)
Отсюда сумма квадратов изменения численности хищника и жертвы – есть величина постоянная. По (3) можно рассчитать изменения численности одного из взаимодействующих видов по изменению численности другого. Например:
ΔN1 = [ R2 – (ΔN2)2]1/2.
Если проанализировать изменения численности хищника и жертвы во времени, можно получить две синусоидальные кривые, максимумы которых сдвинуты относительно друг друга (рис.2), или так называемую «временнэю развертку». При этом на обоих кривых выделяются те же четыре фазы изменения численности взаимодействующих видов, которые выявлены на фазовом портрете системы. Такие циклы в модели Лотки – Вольтерры принято
называть устойчивыми предельными циклами.
Модель Лотки – Вольтерры объясняет циклические колебания численности обоих взаимодействующих популяций следующим образом. Когда численность популяции рыси незначительна, популяция зайца растет очень быстро. Однако увеличение ее численности приводит к возрастанию количества пищи для рысей, поэтому численность последних также начинает возрастать. В свою очередь, увеличение численности рысей приводило к тому, количество потребляемых ими зайцев возрастало, что приводило к резкому снижению числа
последних; затем цикл вновь повторялся. Таким образом, численность хищника и жертвы образуют регулярные циклические колебания. Регуляция численности обоих популяций осуществляется по принципу отрицательной обратной связи.
42. Паразитизм и его значение в экосистемах.
Паразитизм (+ : -) – взаимодействие, при котором один вид (паразит) использует поверхность или внутренние части тела (органы, ткани, клетки) другого организма (хозяина) в качестве среды своего обитания и (или) источника пищи. При этом паразит наносит хозяину определенный вред, часто весьма значительный, вплоть до гибели. Среди хозяев выделяются основные, в которых паразит достигает половой зрелости и производит потомство, и промежуточные, в которых паразит проходит промежуточные стадии жизненного цикла. Часто промежуточных хозяев может быть несколько, иногда их нет вовсе. Как правило, отношения «паразит – хозяин» отличаются определенной видоспецифичностью. Большинство видов паразитов способно паразитировать на определенном виде хозяев или на небольшой группе близкородственных видов. Организм хозяина можно рассматривать как остров, население которого составляют паразиты. Возбудители заболевания могут переселяться с одного на другой только с помощью переносчиков. Чем дальше такие острова друг от друга, тем ниже вероятность передачи инфекции.
Паразиты подразделяются на две основные группы:
Микропаразиты размножаются непосредственно внутри тела хозяина, обычно внутри его клеток. К ним относятся протисты, бактерии и вирусы. Многие из них являются переносчиками опасных заболеваний.
Макропаразиты развиваются на поверхности или во внутренних полостях тела организмов. В их жизненном цикле имеются особые стадии, покидающие хозяев, чтобы заселить новых хозяев. К макропаразитам относятся плоские и круглые черви, членистоногие – вши, блохи и др.
Паразитизм очень широко распространен в биосфере, он встречается во всех царствах живых организмов. Кроме того, все вирусы являются паразитами. Многие типы целиком состоят из паразитов, например, скребни. В значительно меньшей степени паразитизм распространен среди растений, хотя паразитические растения не так уж редки, например омела, раффлезия. Полностью отсутствуют паразиты среди вторичноротых животных.
43.Внутривидовая конкуренция и компенсационный рост.
Если конкуренты принадлежат к одному виду, то взаимоотношения между ними называют внутривидовой конкуренцией. Конкуренция между особями одного вида является наиболее острой и жесткой в природе, поскольку они имеют одинаковые потребности в экологических факторах. Внутривидовую конкуренцию можно наблюдать в колониях пингвинов, где происходит борьба за жизненное пространство. Каждая особь удерживает свой участок территории и агрессивно настроена по отношению к соседям. Это приводит к четкому разделению территории внутри популяции.
Внутривидовая конкуренция на том или ином этапе существования вида встречается почти всегда, поэтому в процессе эволюции у организмов выработались приспособления, снижающие ее интенсивность. Наиболее важные из них — способность к расселению потомков и охрана границ индивидуального участка (территориальность), когда животное защищает место своего гнездовья или определенный участок. Так, в период размножения птиц самец охраняет определенную территорию, на которую, кроме своей самки, не допускает ни одной особи своего вида. Такую же картину можно наблюдать и у некоторых рыб.
Компенсационный рост. Особи могут отставать в росте под воздействием различных факторов – высокой плотности, недостатка пищи, неблагоприятной температуры, воздействия метаболитов и т. п. Снятие лимитирующего фактора часто приводит к резкому ускорению роста аутсайдеров, что определяется как компенсационный рост. Его количественным показателем (Ксоmp) является отношение приростов массы их тела после (ΔW2) и до снятия (ΔW1) лимитирующего фактора, т. е. Ксоmp = ΔW2/ΔW1. Количественные аспекты компенсационного роста исследованы у легочного моллюска – большого прудовика Lymnaea stagnalis из двух водоемов (Голубев, Рощина, 2005). Новорожденные особи до возраста 74–84
суток содержались группами при плотностях от 5,7 до 28,5 экз·л-1. Затем из каждой группы отбирали выборки по 5 самых крупных (лидеры) и самых мелких (аутсайдеры) особей, которых выращивали раздельно при минимальной плотности 2,5 экз л-1 до возраста 150–160 суток. Значения Ксоmp для каждой выборки представлены на рис.8.
У особей из обоих водоемов Кcomp быстро снижаются с увеличением массы их тела при рассадке (Wi). Зависимость между Кcomp и Wi следует уравнению Кcomp = А – blgWi, где А и b – эмпирические коэффициенты. В численной форме данное уравнение для моллюсков из Крутыни имеет вид:
Кcomp = 19,33 – 6,06 lgWi,
а из Припяти:
Kcomp = 30,14 – 10,19 lgWi.
Таким образом, у L. stagnalis самым интенсивным компенсационным ростом обладают наиболее отставшие в росте особи. Чем ниже масса особей при снятии ограничивающего фактора, тем с большей скоростью происходит их компенсационный рост – «эффект сжатой пружины». С увеличением массы особей их способность к компенсационному росту постепенно утрачивается, а у половозрелых особей, достигших размеров, близких к
предельным, вообще отсутствует. Из приведенных уравнений возможно определить значения массы тела, выше которой моллюски полностью теряют способность к компенсационному росту, т.е. Wi, при которой Кcomp равно единице. Для особей из Припяти и Крутыни эти значения равны соответственно 725 и 1060 мг. Это приблизительно соответствует массе, при которой L. stagnalis достигает половой зрелости.
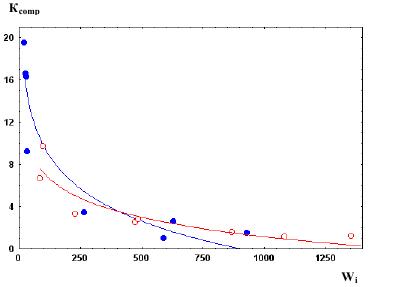
Рис.8. Зависимость между значениями Кcomp у выборок Lymnaea stagnalis из рек Припять (●) и Крутыня (○) от их средней массы (Wi, мг) при рассадке на группы.
В период компенсационного роста генеративный прирост особей (Рg) возрастает прямо пропорционально их соматическому приросту (Ps). При этом значения Pg у аутсайдеров оказались более высокими, чем у лидеров (рис.9). Связь между Pg и Ps следует уравнению Pg = -3,68 + 1,07·Ps.
Рg , мг сутки-1
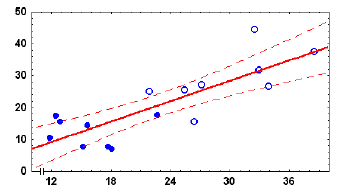
Рис. .9. Зависимость между среднесуточными величинами генеративного (Рg) и соматического (Ps) приростов у лидеров (●) и аутсайдеров (○) Lymnaea stagnalis при компенсационном росте. Пунктир – уровни достоверности 95 %
Таким образом, ускорение соматического роста аутсайдеров при компенсационном росте сопровождается параллельной интенсификацией их размножения. Благодаря способности к компенсационному росту аутсайдеры не являются популяционным балластом, при наступлении благоприятных условий они способны быстро достичь половой зрелости и внести значительный вклад в процесс воспроизводства популяции.
44.Внутривидовая конкуренция у животных. Ее математическая модель.
Внутривидовая конкуренция у животных. Конкурентные отношения в каждой популяции животных по-своему уникальны, тем не менее, существуют их некоторые общие закономерности. Количественным показателем их интенсивности является коэффициент конкуренции (Кс):
Кс = lg(No/Ni), (5)
где No – значение исследуемого параметра при отсутствии конкуренции, Ni – его значение при наличии конкуренции (Бигон и др., 1989). Если увеличение интенсивности внутривидовой конкуренции приводит к снижению исследуемых показателей, значения Кс для них, рассчитанные согласно (5), будут возрастать.
Для естественных популяций ограничивающим ресурсом часто становится пространство. Поэтому повышение плотности популяции, т.е. уменьшение количества пространства, приходящегося на одну особь в ней, всегда приводит к возрастанию внутривидовой конкуренции. Воздействие роста внутривидовой конкуренции, вызванной повышением плотности, на отдельные биологические параметры может быть неодинаковым. Для иллюстрации приведем данные, полученные при выращивании групп особей тепловодного легочного моллюска физелллы Physella integra от рождения и до возраста 32 суток в градиенте плотности от 50 до 2000 экз м-2 (табл. 1). По ним согласно (5) рассчитаны значения Кс для ряда показателей роста и размножения (рис. 4). В качестве No использованы значения соответствующих показателей при минимальной плотности. С увеличением плотности отмечено возрастание вариабельности особей
по массе тела, которую можно оценить по коэффициенту вариации (C.V.):
С.V. = (σ/Wm)·100 %,
где σ – стандартное отклонение (варианса) для массы тела особей в исследованной выборке, Wm – средняя масса особей. При каждой плотности выделялись небольшие группы особей (до 5–10 % от общей численности), отличающиеся резко повышенными или пониженными размерами, по сравнению с основной группой особей. Эти крайние группы можно условно назвать лидерами и аутсайдерами, а основную группу особей – середняками.
Лидерами считались те особи, масса которых превосходит значение Wm+σ, а аутсайдерами – те, масса которых ниже Wm–σ. Табл. 1. Воздействие плотности посадки на некоторые параметры роста и размножения у Physella integra в полевом эксперименте при средней температуре 30оС (по Голубеву, Нагорской, 1997)*
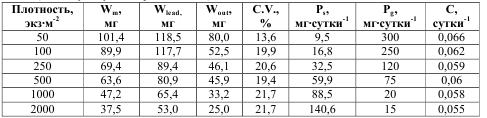
* условные обозначения: Wm – средняя масса особей; Wlead – средняя масса лидеров;
Wout – средняя масса аутсайдеров; C.V. – коэффициент вариации по массе тела; Ps –
среднесуточная соматическая продукция; Pg – среднесуточная генеративная продукция;
C – среднесуточный Р/B-коэффициент для соматической продукции.
Интенсификация внутривидовой конкуренции оказывает большее отрицательное воздействие на рост середняков и особенно аутсайдеров, чем на лидеров, что сказывается соответствующим образом на изменениях значений Кс для этих групп в градиенте плотности (рис.5). Поэтому аутсайдеры находятся под более сильным прессом конкуренции, чем лидеры,
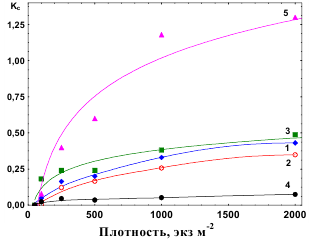
Рис.5. Изменчивость коэффициентов конкуренции (Кс) для различных параметров у Physella integra. 1 – Wm; 2 – Wlead ; 3 – Wout ; 4 – C; 5 – Pg
Аналогичные эксперименты выполнены на головастиках лягушки Rana sylvatica (Wilbur, Collins, 1973). При низких плотностях гистограмма распределения особей по массе тела является вполне симметричной (рис.6).
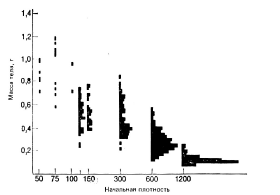
Рис. 6. Гистограммы распределения по массе тела у головастиков лесной лягушки
Rana sylvatica в возрасте 50 суток, выращенных при разной начальной плотности.
Рост головастиков, находившихся в аквариуме при низкой плотности, замедлялся, если в него добавляли воду из аквариума, где головастиков содержали при высокой плотности. Очевидно, увеличение размерной дифференциации в одновозрастных группах растений и животных происходит по механизму положительной обратной связи.
У других организмов регуляторные механизмы могут быть еще более сложными. Например, у гигантских тропических креветок Macrobrachium rosenbergii, выращиваемых в проточных прудах, где метаболиты не накапливались, быстро выделялась небольшая группа самцов-лидеров, отличавшихся очень быстрым ростом. Остальные креветки росли медленно, но если лидеров удаляли, то из них быстро выделалась другая подобная небольшая группа особей и структура популяции быстро восстанавливалась (Ra’anan, Cohen, 1984). Поэтому периодическое изъятие лидеров в процессе культивирования креветок позволяет получить больший выход товарной продукции, чем однократный вылов всей популяции в конце сезона выращивания. У аквариумных рыбок замедление роста наблюдалось даже при их одиночном содержании, если в аквариуме были установлены зеркала, в которых рыбка могла видеть свое отражение.
45.Внутривидовая конкуренция у растений. Ее математическая модель.
Внутривидовая конкуренция у растений. Для популяций многих видов растений, особенно злаковых, установлено, что в широком диапазоне плотностей произведение средней плотности популяции (N) на среднюю массу одного растения (W) является приблизительно постоянной величиной, т. е. N·W = В, (1)
где В – биомасса популяции растений, или их урожай. Поэтому такая закономерность получила название закона постоянного конечного урожая. В таком случае W = В·N-1, или lgW = lgB – lgN. (2)
Отсюда логарифм средней массы одного растения снижается прямо пропорционально возрастанию логарифма плотности популяции. Поэтому угловой коэффициент линии регрессии уравнения (2), характеризующий угол ее наклона, равен –1. Некоторые примеры, иллюстрирующие закон постоянства конечного урожая, представлены на рис.1.
Общая биомасса в посевах клевера подземного Trifolium sunterraneum остается постоянной в широком диапазоне плотности посадки (рис. 1А). У однолетнего растения вульпии Vulpia fasciculata, произрастающего на песчаных дюнах, наклон линии регрессии уравнения (2) с возрастом популяции в конечном итоге приближается к -1 (рис. 1Б, В).
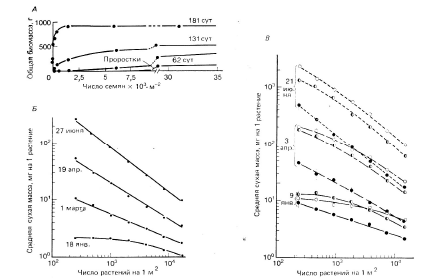
Рис. 1. Закон постоянства конечного урожая при разной плотности посадки в
разновозрастных популяциях некоторых видов растений. А. Изменение биомассы
популяций клевера подземного Trifolium sunterraneum. Б, В. Изменение средней
массы одного экземпляра вульпии Vulpia fasciculatа (из Бигон и др., 1989)
Закон постоянства урожая наблюдается и у некоторых видов животных. Так, c повышением плотности популяции морского брюхоногого моллюска – блюдечка Patella cochlear снижается средний размер особей; однако биомасса популяции остается постоянной в широком градиенте плотности (рис. 2).
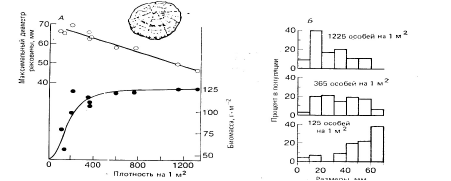
Рис. 2. А. Зависимость среднего размера особей и биомассы популяции Patella
cochlear от ее плотности. Б. Структура микропопуляций блюдечка при разных
плотностях.
С другой стороны, в загущенных посевах многих видов растений, особенно имеющих развитую крону, часто наблюдается самоизреживание (рис.3), т. е. постепенное снижение по мере роста численности экземпляров на единице площади. Это обусловлено тем, что более низкие экземпляры, оказавшись в тени крон более высоких соседних растений, еще более отстают в росте и в конечном итоге погибают. В данном случае регуляция численности популяции происходит по принципу положительной обратной связи. Самоизреживание наиболее выражено в популяциях древесных растений, например, в искусственных лесопосадках ели или сосны. Для злаков самоизреживание менее характерно, особенно при достаточно низких плотностях, поскольку их колосья дают сравнительно мало тени. Для иллюстрации приведем данные экспериментов с плевелом многолетним (райграсом) Lolium perenne (Lonsdale, Watkinson, 1982). Растения высевали при пяти плотностях от 103 до 105 семян на 1 м2 на участках с разным затенением, а затем через 14, 35, 76, 104 и 146 суток после появления проростков на каждом участке определяли среднюю массу растений. При высокой освещенности (рис. 3А) наибольшее самоизреживание отмечено при двух наивысших плотностях, где оно было заметным уже после 35 суток выращивания. При минимальной плотности снижение численности когорты отмечено лишь к концу эксперимента. Снижение плотности когорты (N) приводило к увеличению средней массы одного растения (W). Траектории изменений W при любой плотности когорты направлены к единой прямой (линия самоизреживания), соответствующей уравнению:
W = cN-3/2, (3) где с – константа, равная массе растения при плотности, равной единице.
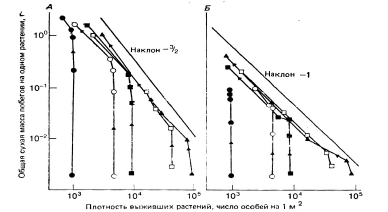
Рис.3. Изменения численности и средней массы растений в когортах плевела
многолетнего Lolium perenne, высаженного при пяти значениях плотности семян на
1 м2: ● – 1000; ○ – 5000; ■ – 10 000; □ – 50 000; ▲ – 100 000 на участках без
затенения (А) и затененном на 83% (Б). Стрелки на линиях для каждой плотности
указывают направление траекторий изменения численности когорты и средней
массы растений в ней по направлению к линии изреживания Поскольку показатель степени в (3) равен –3/2, выявленная закономерность, отмеченная также и для многих других видов растений (рис.4), часто называется «законом трех вторых». Из него следует, что в самоизреживающейся популяции средняя масса одного растения возрастает быстрее, чем снижается их плотность.
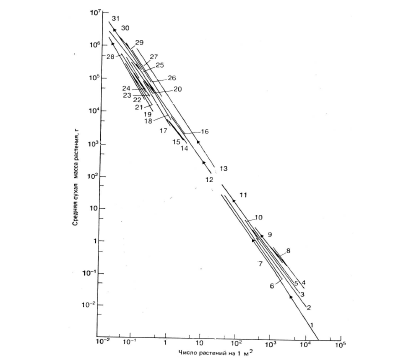
Рис.4. Самоизреживание среди разных видов древесных и травянистых растений. Каждая линия представляет данные для отдельного вида, стрелки на них
указывают направления к общей линии изреживания, которая имеет наклон, близкий к -3/2 (по White, 1980)
При сильном затенении траектории изменения плотности и массы растений направлены к линии регрессии, имеющей наклон, равный –1 (рис.3Б), т.е. в данных условиях выполняется закон постоянства урожая.
Наиболее вероятное объяснение «закона трех вторых» заключается в следующем. Когорта растений представляет массу или объем (параметры, пропорциональные их размеру в степени 3), размещенные под улавливающей свет листовой поверхностью, имеющую некоторую площадь (параметр, пропорциональный размеру в степени 2). В растущей когорте растений индекс листовой поверхности (ИЛП, отношение суммарной площади листьев к площади поверхности почвы, покрываемой кроной) быстро перестает зависеть от общей биомассы когорты и стабилизируется на определенном уровне. Поэтому ИЛП снижается пропорционально росту ее биомассы в степени –3/2 (Бигон др., 1989). При достаточно низких ИПЛ, характерных для начального периода роста растений, чистая первичная продукция их популяций, т. е. урожай, также будет прямо пропорциональной ИПЛ, чем и обусловлен показатель степени в уравнении (2).
Умножив обе части уравнения (3) на N, получаем
W·N = cN-3/2·N.
Поскольку произведение WN представляет биомассу когорты (В), имеем
B = cN-1/2 . (4)
Отсюда
lgB = lgc – 0,5N
Исходя из (4), общая биомасса когорты растений (или их урожай) с понижением плотности их популяции будет возрастать. Этот рост, конечно, не может продолжаться бесконечно. При плотностях популяции ниже определенного предела «закон трех вторых нарушается» и в силу вступает «закон постоянства урожая».
46.Аменсализм и его значение в экосистемах. Математическая модель аменсализма.
Аменсализм (0 : –). К этому типу относятся межвидовые отношения самой разной природы, проявляющиеся в том, что рост численности популяции одного вида взаимодействующей пары в биоценозе оказывает отрицательное воздействие на рост численности популяции другого, но сама не испытывает ответного существенного влияния. Вид, испытывающий в данной паре отрицательное воздействие, называется аменсалом. Односторонние отрицательные взаимодействия известны как между парами видов из различных царств живых организмов – бактерий, грибов, растений и животных, так и между видами из одного царства.
Из взаимодействий между растениями наиболее широко известна конкуренция за свет, воду и биогенные элементы (азот, фосфор, калий, микроэлементы и т. д.), однако эти отношения в большинстве случаев носят характер внутривидовой или межвидовой конкуренции и будут рассмотрены ниже. Наряду с этим, среди взаимодействующих видов растений можно выделить пары видов, один из которых сильно подавляет рост численности другого, не испытывая с его стороны сколько-нибудь заметного ответного влияния. Чаще всего в подобных случаях имеет место аллелопатическое воздействие первого вида на второй посредством выделения в окружающую среду специфических органических ингибиторов, которые тормозят рост или препятствуют размножению и развитию других растений.
Известным примером этого являются виды растений из рода орех (Iuglans), в листьях, скорлупе и корнях которых содержится юглон (5-окси-1,4-нафтохинон). Это вещество из корней, падающих на землю листьев и скорлупы вымывается осадками в почву, где подавляет рост многих видов растений.
Аменсализм между микроорганизмами тесно связан с выделением ими специфических веществ – антибиотиков, отрицательно воздействующих на другие микроорганизмы (антибиоз). В случае, когда продуцент антибиотика не испытывает ответного отрицательного воздействия со стороны ингибируемого вида, можно с уверенностью утверждать о наличии аменсализма.
Отношения между растениями и аменсалами-микроорганизмами связаны с выделением растениями антимикробных токсинов (бактерицидов, фунгицидов и т. п.). Интересным примером этого является возрастающее ингибирование травянистой растительностью деятельности почвенных нитрифицирующих бактерий в процессе сукцессии наземных экосистем. Этим достигается сохранение связанного азота в почве, как важнейшего фактора питания растений. Не менее широко распространены и случаи, когда аменсалами являются растения, рост и развитие которых подавляют токсины, выделяемые микроорганизмами или грибками. Например, различные виды грибков рода Fusarium продуцируют соединения, вызывающие у растений нарушение водного обмена.
Аменсализм между прокариотами и животными ярко иллюстрирует цветение водоемов, когда поступающие в воду токсины, выделяемые размножающимися или отмирающими планктонными цианобактериями, приводят к массовой гибели многих видов водных беспозвоночных и рыб. Интересным примером аменсализма между животными и грибами является муравей-листорез Atta sexdens, который «выращивает» в своих гнездах грибы, используемые затем в пищу. Муравьи в особых железах вырабатывают и постоянно разбрызгивают по гнезду фенилуксусную кислоту и миркамицин. Первое из этих веществ ингибирует рост бактерий, а второе – прорастание спор тех видов грибов, которые не пригодны для своего питания. Типичным примером аменсализма между животными и растениями является вытаптывание стадами крупных растительноядных млекопитающих (слоны, копытные) травяного покрова. В результате ухудшаются условия существования не только травянистых растений, но и многих видов животных, обитающих на поверхности почвы или в почвенном покрове.
Математическая модель аменсализма. Один из способов количественного описания отношений аменсализма между двумя видами заключается в модификации логистического уравнения роста численности взаимодействующих популяций. Приведем данную модель в изложении В. Д. Федорова и Т. Г. Гильманова (1980). Допустим, между популяциями двух видов существуют отношения аменсализма. Тогда скорость роста численности одной из них выражается уравнением
dN1/dt = r1N1(1 – N1/K1), (1)
где r1 – максимально возможная удельная скорость роста популяции первого вида, достигаемая при ее минимальной плотности; К1 – ее предельная (максимально возможная) численность, что может являться показателем емкости для популяции нее. Предположим, что с увеличением численности первого вида (N1) уменьшается емкость среды для второго вида, т.е. аменсала. В простейшем случае это уменьшение можно выразить линейным уравнением:
К2(N1) = K2 – a21N1,
где К2(N1) – предельная численность популяции вида-аменсала, когда численность популяции первого вида достигла N1; K2 – предельная численность популяции вида-аменсала в отсутствии первого вида; a21 (читается «а два-один») – коэффициент ингибирования, показывающий, на сколько особей снижается предельная численность вида-аменсала при росте численности популяции первого вида (N1) на одну особь. Очевидно, что
a21 = [К2 – K2(N1)]/N1. (2)
Тогда скорость роста численности популяции вида-аменсала (N2) в зависимости от достигнутой численности популяции первого вида (N1) описывается уравнением:
dN2/dt = r2N2[1 – N2/(K2 – a21N1)]. (3)
Определим условия, при которых скорость роста популяции вида-аменсала, заданная (3), становится равной нулю, т.е., когда r2N2[1 – N2/(K2 – a21N1)] = 0. (4)
Поскольку значения r2 и N2 в (.4) явно не равны нулю, остается предполагать, что 1 – N2/(K2 – a21N1) = 0. Отсюда получим зависимость численности популяции вида-аменсала (N2) от численности популяции вида, взаимодействующего с ним (N1):
N2 = K2 – a21N1. (5)
Линия регрессии уравнения (16.5) на графике представляет прямую линию, или изоклину (рис.1), которая является геометрическим местом точек, представляющих такие комбинации N1 и N2, при которых удельная скорость роста популяции вида-аменсала (r2) равна нулю.
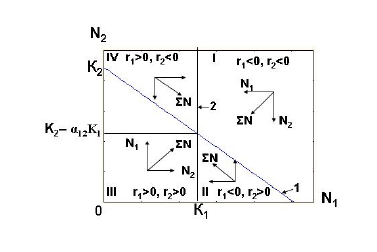
Рис. 1. Фазовый портрет двухвидовой системы с аменсализмом при слабом ингибировании. 1 – линия регрессии уравнения (16.5), отражающая снижение предельной численности популяции вида-аменсала с увеличением численности
популяции первого вида. 2 – линия, отражающая постоянство предельной численности популяции вида, взаимодействующего с аменсалом.
Легко видеть, что значение a21 соответствует тангенсу угла наклона изоклины (16.4) к оси абсцисс. Внизу и слева от изоклины при всех комбинациях N1 и N2 значения r2 для популяции вида-аменсала положительны, а сверху и справа от нее – отрицательны.
Аналогичным образом, приравняв к нулю (рис.1), легко убедиться, что график зависимости численности первого вида от численности вида-аменсала представляет перпендикуляр к оси абсцисс, в точке, соответствующей предельной численности первого вида, т. е. К1. Это является графической иллюстрацией положения, что изменения численности популяции вида-аменсала не оказывают никакого влияния на численность вида, взаимодействующего с ним. Справа от перпендикуляра скорость роста популяции первого вида (r1) отрицательна, а слева – положительна.
Пересечение двух прямых на графике (рис. 1) делит плоскость графика (N2 – N1) на четыре сектора (I – IV). Определим суммарные значения изменения численности обеих взаимодействующих популяций в каждом из них. В первом секторе численность популяций первого и второго видов (N1 и N2) снижается, что показано соответствующими векторами N1 и N2. Суммарную численность обеих взаимодействующих популяций (ΣN), которая также будет снижаться, можно определить по правилу сложения векторов. Суммарный вектор ΣN направлен к точке пересечения прямых на рис. 1. Аналогичным образом можно найти направления векторов ΣN в остальных секторах и убедиться, что все они также направлены к точке пересечения. Таким образом, несложный анализ системы уравнений (1) и (3) показывает, что при положительных значениях коэффициентов r1, r2, K1, K2 и a21 исход отношений между взаимодействующими видами определяется соотношениями этих коэффициентов. Существует два варианта этого исхода. В первом варианте, когда K2/K1 > a21 (слабое ингибирование) точка пересечения прямых лежит выше оси абсцисс. В этом случае возможно
единственное устойчивое состояние, при котором сосуществуют обе взаимодействующие популяции. К нему сходятся решения из любого начального состояния с положительными значениями численности обеих популяций. Нетрудно найти, что предельная численность популяции вида-аменсала снизится до уровня, равного К2 – а21К1, а первого вида, взаимодействующего с ним, – останется прежней, т. е. равной К1. Это означает, что оба взаимодействующих вида могут сосуществовать, хотя максимальная численность вида-аменсала снизится по сравнению с таковой при отсутствии взаимодействия.
Во втором случае, при K2/K1 < a21 (сильное ингибирование), точка пересечения прямых находится ниже оси абсцисс (рис. 2). В этом случае система также имеет единственное устойчивое состояние, при котором, однако, значение К2 – а21К1 является отрицательным. Из этого следует, что численность популяции вида-аменсала с увеличением численности популяции первого вида постепенно снизится до нуля, т. е. произойдет его полное вытеснение.
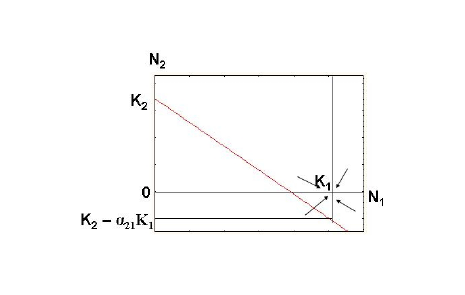
Рис. 2. Фазовый портрет двухвидовой системы при аменсализме с сильнымингибированием.
Результаты наблюдений над лабораторными культурами бактерий кишечной палочки Escherichia coli, клебсиеллы Klebsiella sp. и схигеллы Shigella flexneri показывают примеры реализации обоих вариантов. Схигелла в паре с клебсиеллой вполне может существовать, хотя максимальная плотность ее популяции в смешанной культуре снижается почти на три порядка по сравнению с монокультурой. Напротив, при выращивании совместно с кишечной палочкой схигелла почти полностью вытесняется.
47.Протокооперация и мутуализм, их значение в экосистемах и эволюции Биосферы. Математическая модель мутуализма.
Протокооперация (+ : +) возникает, когда оба вида получают пользу от взаимодействия друг с другом, однако это взаимодействие не носит обязательного характера и оба вида могут вполне обойтись без него. Например, некоторые птицы питаются паразитами, живущими на поверхности кожных покровов крупных млекопитающих (гиппопотамы, полорогие и т.д.), очищая тем самых их от паразитов. Однако эти же птицы способны потреблять и другие виды корма, потому они выживают и в отсутствии млекопитающих. Первым шагом к развитию отношений протокооперации между двумя видами, по-видимому, является комменсализм. Начальные стадии протокооперации между различными видами встречаются в природе повсеместно. Так, в морских экосистемах многие виды актиний прикрепляются к спинам крабов (иногда крабы сажают их сами), маскируя и защищая их своими щупальцами со стрекательными клетками. В свою очередь, актинии используют крабов как транспортное средство и потребляют остатки их пищи.
К протокооперации вполне возможно отнести отношения между человеком и такими домашними животными, как кошки и собаки. Эти животные получают от человека кров и пищу, а человек от них – положительные эмоции. Однако, будучи выброшенными на улицу, собаки и кошки способны выжить и самостоятельно. При наличии или отсутствии протокооперации популяционные
характеристики обоих взаимодействующих видов заметным образом не изменяются. К сожалению, количественные аспекты этого протокооперации
исследованы еще очень слабо.
Мутуализм (+ : +) – это взаимодействие между парами видов (мутуалистов), при котором увеличение (или снижение) численности популяции одного вида в конечном итоге приводит к увеличению (или снижению) скорости роста численности популяции другого вида. При мутуализме один или оба вида-мутуалиста не могут существовать друг без друга. По крайней мере, популяционные характеристики одного вида (скорость роста и размножения, выживаемость и пр.) значительно снижаются в отсутствие другого. Мутуализм имеет ряд градаций: факультативный, когда один вид- мутуалист получает пользу от своего партнера, но не зависит от него; облигатный для одного, но факультативный для другого; облигатный для обоих. В последнем случае оба вида тесно связаны физиологически, часто один вид поселяется в теле другого. Например, одноклеточные зеленые водоросли живут в теле гидр, некоторые даже проникают в их клетки. Иногда мутуалистические взаимоотношения неправильно называют симбиозом (от греч. сим – «вместе», биос – «жизнь»). Однако сейчас симбиоз определяется как любая форма взаимодействия совместно проживающих организмов. Поэтому симбионтные взаимоотношения популяций различных видов могут иметь самый разный характер – нейтрализм, комменсализм в форме «квартирантства», протокооперация, паразитизм и собственно мутуализм.
Мутуалистические отношения в природе распространены очень широко, они известны между видами из всех царств органического мира. Они имеют огромное значение роль в функционировании и эволюции экосистем, от них в очень большой мере зависит биологическая продуктивность биосферы. Например, значительная часть продукции лесных экосистем создается благодаря микоризе (от греч. микос – «гриб» и ризос – «корень»), т.е. сообществу бактерий и грибов, поселяющихся на корнях древесных растений. В частности, из приблизительно 3500 видов сосудистых растений флоры бывшего СССР, около 80 % (в их числе все виды голосемянных) имеют ту или иную форму микоризы. Интересно, что все попытки разведения тропических орхидей в европейских оранжереях оказывались безуспешными, пока не было выяснено, что для прорастания семян орхидей и последующего развития проростков необходимо их взаимодействие с определенными видами грибов, которые не способны жить в европейских почвах. В данном случае мы имеем пример облигатного мутуализма орхидей и их грибов-симбионтов. Биологическая фиксация атмосферного азота микроорганизмами, поселяющимися в тканях корней высших растений, является существенным, а зачастую основным источником поступления связанного азота в наземные и водные экосистемы. Растения снабжают бактерии углеводами и другими соединениями, используемые ими для своего существования и фиксации молекулярного азота, который затем усваивается растениями. Чаще всего азотфиксирующие бактерии поселяются в корнях бобовых растений, вызывая на них специфические утолщения – клубеньки; поэтому их называют клубеньковыми бактериями. Благодаря им бобовые растения способны поселяться и расти на почвах, бедных минеральным азотом, что характерно для начальных стадий сукцессии наземных экосистем. Известны и другие примеры мутуализма азотфиксирующих прокариотов и высших растений. В их числе симбиоз разных видов рода ольха (Alnus) c актиномицетами, водного папоротника азоллы Azolla c цианобактерией анабеной Anabaena azollae. Азоллу широко используют в Японии и Индокитае как естественное удобрение для рисовых полей. Большинство видов цветковых растений опыляется насекомыми, при этом само опыление происходит попутно в процессе сбора нектара или пыльцы из цветков. На образование нектара, который, по-видимому, необходим растению только для привлечения опылителей, расходуется большое количество органических веществ; однако эти траты многократно компенсируются пользой от опыления. Многие виды растительноядных животных из самых разных групп (млекопитающие, термиты) усваивают углеводы потребляемой ими пищи с помощью многочисленных симбионтных бактерий и инфузорий, живущих в
их пищеварительном тракте. При этом микроорганизмы получают постоянный источник пищи и довольно стабильные условия существования, а животные –
доступные для переваривания вещества из корма, который они не способы
переработать с помощью собственных ферментов.
Мутуалистическими являются отношения между человеком и культурными растениями и сельскохозяйственными животными – крупным рогатым скотом, лошадьми, свиньями, овцами, козами и пр. Последние за длительный период сосуществования с человеком так изменились, что уже не способны выжить самостоятельно без его помощи. С другой стороны современное человечество также не может прожить без пищевых продуктов, даваемых культурными растениями и домашними животными.
Мутуализм играет огромную роль в эволюции жизни на Земле. Согласно теории симбиогенеза, эукариотические организмы возникли в результате внутриклеточного симбиоза. Предполагается, что в прокариотной анаэробной и гетеротрофной бактерии поселились паразитические аэробные и автотрофные (фотосинтезирующие) бактерии. Эти внутриклеточные паразиты в течение длительной эволюции превратились соответственно в митохондрии и хлоропласты, а сам хозяин – в эукариотический аэробный организм (Маргелис, 1983; Давыденко, Даниленко, 2003). Длительное время лишайники рассматривались как пример симбионтного мутуализма грибов с цианобактериями, зелеными, желто-зелеными или бурыми водорослями. Считалось, что гриб снабжает своих симбионтов водой и минеральными солями, получая взамен созданные при фотосинтезе органические вещества. В действительности эти взаимоотношения скорее имеют характер двустороннего паразитизма, более сильного со стороны гриба, поскольку гифы последнего окружают клетки бактерий или водорослей и могут даже проникать в них. Гриб в качестве объекта питания использует также отмершие клетки своих симбионтов, являясь в данном случае сапротрофом (Трасс, 1977). Этот пример показывает, что грань, отделяющая мутуализм от других типов межвидовых отношений, может быть весьма нечеткой.
Математическая модель мутуализма приведена в изложении В.Д.Федорова и Т.Г Гильманова (1980). При описании динамики популяций, взаимодействующих по типу мутуализма, целесообразно, как и в разобранных выше случаях аменсализма и комменсализма, исходить из уравнения логистического роста. Тогда рост популяции первого вида в отсутствие другого следует уравнению:
dN1/dt = N1r1(1– N1/K1),
а рост популяции второго вида в отсутствие первого – уравнению:
dN2/dt = N2r2(1– N2/K2),
где N1 и N2 – численность первого и второго видов; r1 и 2 – удельные скорости
роста популяций первого и второго видов при их минимальной плотности; K1 и K2 – предельные значения численности популяций первого и второго видов, что может рассматриваться как показатель емкости среды для них. Допустим, взаимное влияние видов друг на друга проявляется в том, что при увеличении численности каждого из них пропорционально увеличивается и емкость среды для другого, например, новых укрытий, мест для прикрепления, гнездования и т.п. Тогда численность популяции первого вида будет расти согласно:
dN1/dt = N1r1[(1– N1/(K1 + χ21N1)], (8)
где χ21 – «коэффициент мутуализма», показывающий, насколько увеличивается
емкость среды для первого вида при увеличении численности популяции второго вида на одну единицу. Уравнение для роста численности популяции второго взаимодействующего вида имеет вид, аналогичный (8):
dN2dt = N2r2[(1– N2/(K2 + χ12N2)], (9)
где χ12 – «коэффициент мутуализма», смысл которого также аналогичен
приведенному выше.
Допустим также, что значения емкости среды для каждого вида – К1(N2) и К2(N1) линейно возрастают с увеличением численности другого, т.е. К1(N2) = K1 + χ21N2 и К2(N1) = K2 + χ12N1. Способом, аналогичным описанным выше, построим графики зависимости численности каждого из видов-мутуалистов от другого взаимодействующего вида (рис. 6).

Рис. 6. Фазовый портрет двухвидовой системы при мутуализме. 1 и 2 – рост
предельной численности популяций первого и второго видов Анализ этой модели приводит к заключению, что при положительных значениях входящих в систему уравнений (8) и (9) параметров r1, r2, K1, K2, χ21 χ12 она имеет единственную устойчивую стационарную точку. Ее координаты, соответствующие максимально возможным значениям численности популяций обоих взаимодействующих видов, равны
N1max = K1 + χ12K2 и N2max = K2+ χ21K1.
В случае, когда χ12 = χ21, линии 1 и 2 на рис. 16.6. будут располагаться параллельно друг другу; отсюда численность обеих взаимодействующих популяций может расти бесконечно. Однако подобный случай не имеет15 биологического смысла, поскольку этот рост остановят другие лимитирующие
факторы среды.
Иллюстрацией этой модели являются эксперименты с двумя видами бактерий – одного из штаммов молочной палочки Lactobacterium arabinosus и стрептококка Streptococcus faecalis. Указанный штамм не способен синтезировать фенилаланин, который продуцирует стрептококк. В свою очередь, последний нуждается в фолиевой кислоте, которую производит молочная палочка. Поэтому при раздельном культивировании на средах, не содержащих этих соединений, оба вида растут очень плохо. Однако при совместном культивировании каждый вид обогащает среду соединением, необходимым для другого, в результате чего смешанная культура дает интенсивный S-образный рост (рис. 5б).
48.Комменсализм, его значение в экосистемах. Математическая модель комменсализма.
Комменсализм (+ : 0). К этой категории относятся разнообразные случаи отношений между двумя видами, когда первый вид, называемый «хозяином», положительно воздействует на второй вид, называемый «комменсалом», но не испытывает никакого ответного влияния с его стороны. Наиболее распространенная форма комменсализма (иногда ее не совсем правильно называют «нахлебничество»), когда вид-комменсал тем или иным способом получает от вида-хозяина пищу, чем обеспечивает свое существование. Так, многие мелкие виды животных поедают остатки пищи, экскременты или прижизненные выделения более крупных животных.
Кроме того, положительное влияние хозяина на комменсала может заключаться в предоставлении жилища («квартирантство»), укрытия от хищников, субстрата для прикрепления.
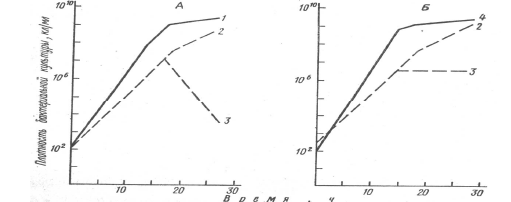
Рис. 3. Аменсализм между бактериями Escherichia coli, Klebsiella sp. и Shigella flexneri. Рост E .coli (1) и Klebsiella sp. (4) в чистой и в смешанной культуре с Sh.
flexneri. 2 и 3. Рост Sh. flexneri соответственно в чистой и смешанной культуре Наконец, хозяин может способствовать распространению и размножению комменсала («форезия»), выполняя роль переносчика его взрослых особей или молоди. Отношения типа комменсализма встречаются преимущественно среди
животных, растений и микроорганизмов. При этом растения, будучи автотрофами, обычно выступают в роли хозяев. Например, существенную роль в круговороте органического вещества в биогеоценозах играют микроорганизмы, живущие на поверхности листьев (в филлосфере) и в прикорневой зоне (в ризосфере) растений, которые перерабатывают их прижизненные выделения. Те микроорганизмы, которые не оказывают существенного положительного влияния на растение-хозяина, например, не снабжают его соединениями азота или другими веществами, вполне могут
рассматриваться как комменсалы. Комменсализм животных по отношению к растениям также широко распространен и имеет самые разнообразные формы – от использования в пищу живых или мертвых тканей опавшей листвы до квартирантства и форезии. Примером последнего случая является расселение животных через водные преграды на «плотах», т. е. на плавающих стволах или ветвях деревьев. Многочисленны случаи комменсализма среди животных разных видов. Например, у льва – самого крупного хищника в биоценозе африканской
саванны имеется целый ряд комменсалов, питающихся его объедками: гиены, шакалы, птицы-падальщики и др. В морских биоценозах аналогичным примером являются стайки мелких рыб разных видов, следующие за акулой.
В норах млекопитающих, птичьих гнездах, муравейниках и жилищах других животных обитает множество «квартирантов», извлекающих из этого соседства большую выгоду, но нейтральных по отношению к хозяевам. У водных организмов, рыб, моллюсков, китообразных имеются многочисленные комменсалы, живущие на поверхности тела или во внутренних полостях своих хозяев. Например, в мантийной полости устриц обитают небольшие крабы, однако они иногда нарушают статус «гостя» и питаются тканями тела хозяина. В отдельных случаях хозяин даже может получать от своего сожителя некоторую пользу. Поэтому комменсализм в случае «квартирантства» иногда приближается к паразитизму или протокооперации. Среди комменсалов заметное место занимают и микроорганизмы, главным образом, бактерии, обитающие на внешних покровах или в пищеварительном тракте животных.
В зависимости от способности комменсала существовать самостоятельно, не пользуясь услугами хозяина, различают факультативный и облигатный комменсализм. Так, гиена и шакал по отношению ко льву являются факультативными комменсалами, поскольку они способны самостоятельно добывать себе корм. Напротив, птицы-падальщики (например, грифы) практически потеряли способность к самостоятельной охоте и потому являются облигатными комменсалами. В результате длительной сопряженной эволюции видов, взаимодействующих по типу комменсализма, у комменсала могут
сформироваться специфические морфологические, физиологические и биохимические адаптации, которые повышают его шансы на выживание. Многие комменсалы, связанные со своими хозяевами отношениями форезии,
имеют разнообразные приспособления (щетинки, крючочки, присоски и т. п.), обеспечивающие прочный механический контакт с хозяином. Интересным
примером биохимической адаптации являются некоторые рыбы, выработавшие иммунитет к ядовитым выделениям медуз и актиний, благодаря чему укрываются от хищников между их щупальцами.
Математическая модель комменсализма приведена нами в изложении В.Д.Федорова и Т.Г.Гильманова (1980). Как и в случае аменсализма, для математического описания комменсализма может быть использовано уравнение логистического роста популяции. Однако разница состоит в том, что с возрастанием плотности популяции хозяина емкость среды для комменсала будет не уменьшаться, как при аменсализме, а возрастать. В простейшем случае получаем:
К2(N1) = K2 + β21N1,
где К2 – максимально возможная численность популяции комменсала в отсутствие хозяина, что можно рассматривать как показатель емкости среды
для комменсала; β21 – коэффициент комменсализма, показывающий, насколько
возрастает емкость среды для комменсала (К2(N1)) при увеличении численности
популяции хозяина (N1) на одну единицу. Очевидно, случай, когда значение К2 для комменсала в отсутствие хозяина равно нулю, соответствует облигатному комменсализму, а все случаи, где К2 > 0, – факультативному комменсализму. Значение β21 рассчитывается аналогично (2). Тогда для описания изменения скоростей роста двух популяций, взаимодействующих по типу комменсализма, получаем уравнение для хозяина:
dN1/dt = r1N1(1 – N1/K1) (6)
и комменсала:
dN2/dt = r2N2[1 – N2/(K2 + β21N1)]. (7)
Способом, аналогичным для взаимодействия по типу аменсализма, построим графическую модель изменения численности популяций хозяина и комменсала друг от друга (рис. 4). Исследование системы уравнений (6) и (7) показывает, что при положительных r1, r2, К1, K2 и β21 существует единственное устойчивое
стационарное состояние численности взаимодействующих популяций. При этом предельная численность популяции комменсала возрастет до К2 + β21К1, а численность популяции хозяина останется неизменной, т. е. К1.
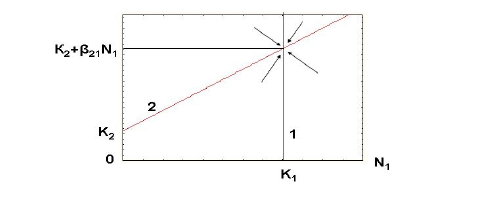
Рис. 4. Фазовый портрет динамики двухвидовой системы при комменсализме.
1. Предельная численность популяции вида-хозяина, которая остается неизменной;
2. Рост предельной численности популяции вида-комменсала при возрастании
численности популяции вида-хозяина В качестве примера пары популяций, взаимодействующих по типу комменсализма, приведем поликультуру из зеленой одноклеточной водоросли хлореллы Chlorella pyrenoidosa и гетеротрофной бактерии Flavobacterium sp., содержащуюся на свету в неорганической питательной среде (рис. 5а). Источником энергии для бактерии являются прижизненные выделения хлореллы, создаваемые ей в процессе фотосинтеза, поэтому в отсутствие хлореллы рост численности Flavobacterium sp. значительно замедляется.
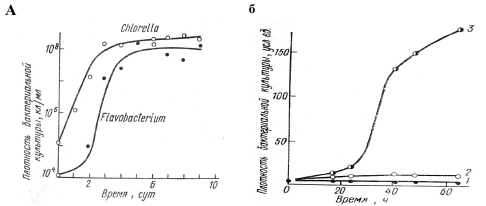
Рис. 5. а. Комменсализм Flavobacterium sp. по отношению к Chlorella pyrenoidosa. б.
Мутуализм между Lactobacterium arabinisus и Streptococcus faecalis. 1 и 2 –
раздельное культивирование обоих видов на синтетической среде, лишенной
фенилаланина и фолиевой кислоты; 3 – суммарная численность обоих видов,
выращиваемых в смешанной культуре
49.Экологическая ниша и межвидовая конкуренция. Принцип конкурентного исключения.
Общие положения. Считается, что термин «экологическая ниша» впервые применил в 1910 г. Р. Джонсон, анализируя распространение видов в зависимости от абиотических и пищевых факторов. Однако его широкое использование в экологической литературе началось с работы американского орнитолога Дж. Гринелла (1917), посвященной экологии калифорнийского пересмешника Tohostoma redivivum. Так, Гринелл указывал, что ниша пересмешника характеризуется целым комплексом специфических условий (абиотические факторы, пища, укрытия и др.), которые имеются лишь в чапарале (сообщество засухоустойчивых кустарников) Калифорнии и Мексики, поэтому пересмешник может обитать только там (Федоров, Гильманов, 1980, Гиляров, 1990). В развитии концепции экологической ниши большой вклад внес английский эколог Ч. Элтон. В своей книге «Экология животных» (1927) он определял нишу животного как «образ жизни и в особенности способ питания в том смысле, в каком говорят о профессии или занятии в человеческом обществе». В других местах Элтон указывал, что «ниша животного означает его место в биотической среде, его отношения с пищей и врагами», подчеркивая, что идея ниши имеет чисто экологический, а не таксономический характер. Так нишу животных, питающихся цветочной пыльцой и попутно опыляющих цветки, в умеренной зоне Евразии занимают насекомые, в тропических лесах Южной и Центральной Америки – наряду с ними птицы (колибри), а в Австралии – даже мелкие сумчатые млекопитающие (кускусы). Подобные виды, занимающие одинаковые нищи в однотипных экосистемах разных географических регионов, называются
экологическими эквивалентами.
Концепция функциональной ниши Элтона («элтонианская ниша»), отождествляющая ее с положением вида в трофической структуре биогеоценоза, оказала значительное влияние на развитие экологии. Именно к ней относится широко известное выражение Ю. Одума «местообитание – это адрес вида, а экологическая нища – его «профессия». Однако чисто качественная трактовка концепции затрудняла ее практическое применение. Напротив, ниша в трактовке Гринелла ассоциируется с градиентами комплекса экологических факторов, в которых существует вид, что допускает ее количественный анализ. Новый этап в развитии концепции экологической ниши начался в 1957 г, когда Дж. Хатчинсон предложил понятие многомерной ниши, которое по существу, явилось дальнейшей разработкой представлений Гринелла. В формализованном подходе Хатчинсона ниша – это «N-мерный гиперобъем, охватывающий весь диапазон условий», в котором особь (или популяция) существует и может успешно воспроизводить себя». Хатчинсон предложил различать фундаментальную нишу, охватывающую весь диапазон условий среды, в котором вид способен существовать, и реализованную нишу, т. е. тот реальный диапазон условий среды, в котором он существует в реальной экосистеме. Как правило, реализованная ниша меньше фундаментальной, поскольку диапазон факторов среды, в которых вид существует в экосистеме, зависит не только от степени его толерантности к ним (эврибионтные и стенобионтные виды), но и от его отношений с другими видами. Например, рачок-бокоплав Gammarus duebeni обитает в солоноватых водах вдоль всего побережья Британских островов; однако в Ирландии он населяет также и внутренние пресные водоемы, поскольку там отсутствует его конкурент – пресноводный Gammarus pulex. Напротив, во внутренних водоемах Великобритании, где доминирует Gammarus pulex, G.duebeni отсутствует (Константинов, 1986). Э. Пианка (1981) предложил принципиально иную трактовку экологической ниши, определив ее как «общую сумму адаптаций организменной единицы» или же как «все разнообразные пути приспособления данной организменной единицы к определенной среде». Такая трактовка отражает способность организма или популяции осваивать среду своего обитания и подразумевает пути ее использования. Она же предлагает ответ на дискуссионный вопрос о существовании в экосистеме «пустующих» экологических ниш: их нет, поскольку каждый новый вид приходит в экосистему с «собственной» нишей, которая уничтожается вместе с исчезновением этого вида из экосистемы. Тем не менее, такой подход к трактовке концепции экологической ниши не получил всеобщего признания.
Межвидовая конкуренция – это любое взаимодействие между двумя или более популяциями, которое отрицательно сказывается на их росте и выживании. Независимо оттого, что лежит в основе межвидовой конкуренции – использование одних и тех же ресурсов, межвидовые химические или алеллопатические взаимодействия или хищничество, - она может привести либо к взаимному приспособлению видов, либо к вытеснению одного вида другим.
Межвидовую конкуренцию, так же как и внутривидовую можно разделить на 2 основных типа — интерференцию и эксплуатацию, хотя в любом конкретном взаимодействии можно выявить элементы и того, и другого типа. В случае эксплуатационной конкуренции особи взаимодействуют друг с другом косвенно, реагируя на количество ресурса, пониженное вследствие активности конкурентов. Пример такого взаимодействия дают эксперименты Тилмана и его коллег на диатомовых водорослях. Когда межвидовая конкуренция основана на использовании общего ресурса, то один вид потребляет этот ресурс и уменьшает его количество до такого уровня, при котором скорость роста, размножения или выживаемость другого вида снижается. Морские желуди, которых изучал Коннелл, напротив, дают нам хороший пример интерференционной конкуренции. Balanus, в частности, прямо и физически препятствует закреплениюChthamalus на ограниченных участках каменистого субстрата.
Другим важным моментом является то, что межвидовая конкуренция (подобно внутривидовой) часто бывает сильно асимметричной, т. е. ее последствия не одинаковы для обоих видов.
Принцип конкурентного исключения
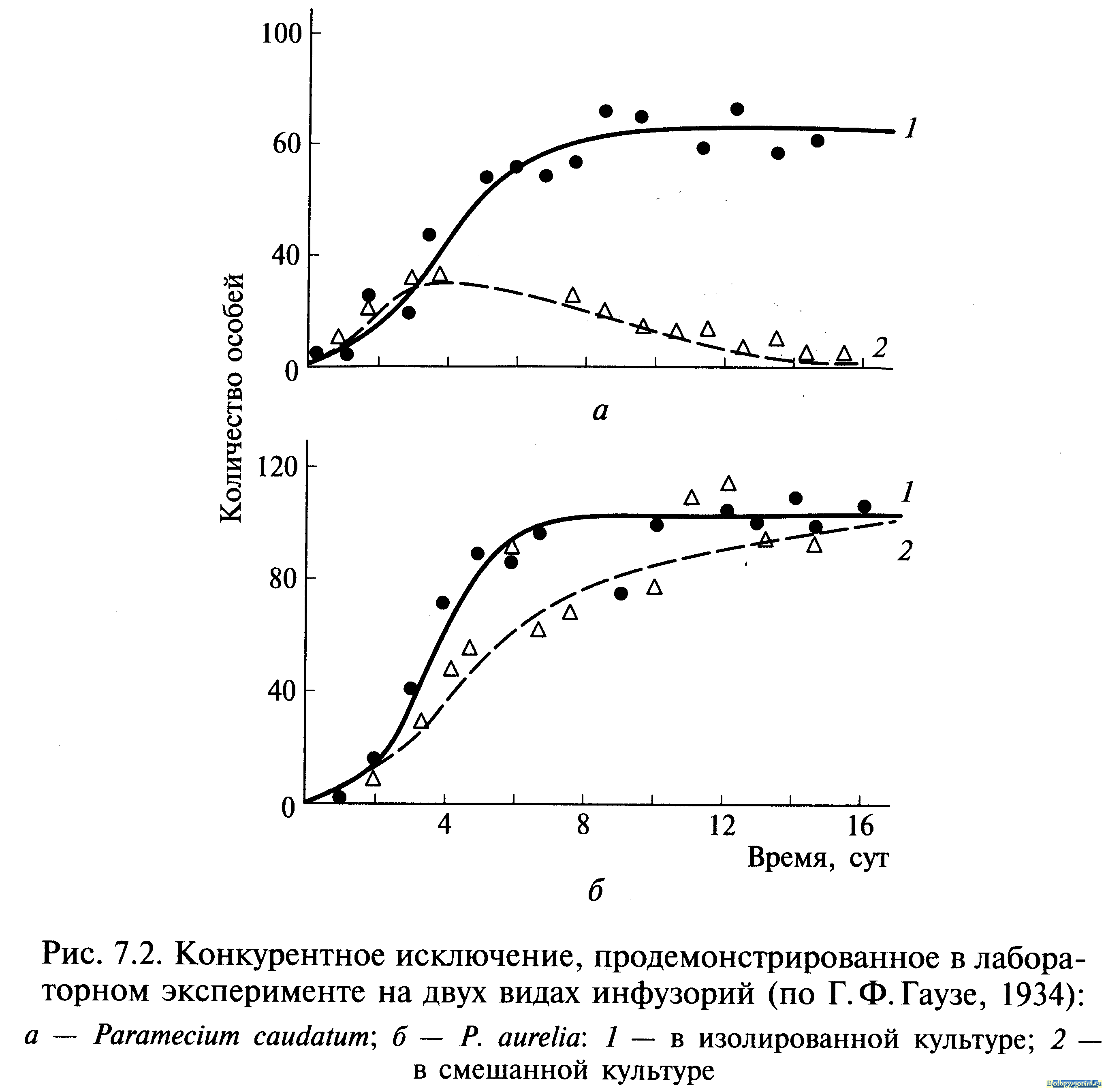
Внутривидовая конкуренция сильнее межвидовой, но правило конкуренции распространяется и на последнюю: конкуренция между двумя видами тем сильнее, чем ближе их потребности. В крайнем случае можно допустить, что два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое-то время обязательно будет вытеснен. Это положение получило статус закона, известного как принцип конкурентного вытеснения (исключения), или принцип Гаузе — в честь Г. Ф. Гаузе, впервые продемонстрировавшего это явление в экспериментах с инфузориями (рис. 7.2). При изолированном содержании численность популяций обоих видов инфузорий увеличивается до тех пор, пока не достигнет максимума. В смешанной культуре популяция второго вида (рис. 7.2, б, кривая 2) растет быстрее, чем первого (рис. 7.2, а, кривая 2), что приводит к ограничению ее численности. Если не произойдет изменений в параметрах конкуренции, то конкурентное исключение (вымирание первой популяции) — лишь вопрос времени. Иногда два близких вида, имеющих одинаковые пищевые потребности, живут на одной территории, не конкурируя друг с другом. Такие, казалось бы, исключения из принципа Гаузе можно обнаружить среди птиц. В Англии большой баклан и хохлатый баклан совместно гнездятся на одних и тех же скалах и кормятся в одних и тех же водах, но они вылавливают разную пищу. Большой баклан ныряет глубоко и питается преимущественно бентосными животными (камбала, креветки), тогда как хохлатый баклан охотится в поверхностных водах на сельдевы рыб. Подобные факты, а их много, подводят нас к понятию экологической ниши, предложенному Ч.Элтоном еще в 1927 г. По выражению Одума, местообитание — это адрес вида, тогда как экологическая ниша — это род его занятий. Иными словами, знание экологической ниши дает возможность ответить на вопросы, где, чем и как питается вид, чьей добычей он сам является, каким образом и где он размножается и т.д. Современное определение звучит следующим образом: экологическая ниша — это роль вида в переносе энергии в экосистеме. Определение точное, но краткое, не раскрывающее в полной мере значение этого термина.
50.Понятия экосистемы и биогеоценоза, структурно-функциональная организация биогеоценоза.
Любая совокупность организмов, существующих в определенном объеме пространства, и их связей между собой и окружающей средой определяется как экологическая система, или экосистема. Термин «экосистема» впервые предложил английский эколог А. Тэнсли в 1935 г., хотя представления об экосистемах как особой форме взаимоотношений организмов со средой своего обитания возникли значительно раньше. Легко видеть, что понятие экосистемы является очень широким и не имеет ранга размерности. Экосистемой можно назвать любой объем пространства, в котором существуют живые организмы, хотя бы один. Ей может быть и капля
воды с одной амебой, пруд, лесной массив, пилотируемый космический корабль и вся Биосфера Земли.
В 1944 г. русский геоботаник В. Н. Сукачев предложил более конкретный термин «биогеоценоз». Он определяется как участок земной поверхности (в том числе и водной) с определенной совокупностью живых организмов и вполне однородными условиями среды, достаточно отделенный от других участков, в пределах которого осуществляется более-менее замкнутый процесс круговорота веществ. Примерами биогеоценозов являются пруд, луг, лесной массив, болото и т. д.
Термины «экосистема» и «биогеоценоз» достаточно близки, но не полностью тождественны, поскольку понятие биогеоценоза значительно более конкретно, чем экосистемы. Например, капля воды и Биосфера являются экосистемами, но не биогеоценозами. Однако поскольку термин «экосистема» легче произносить, в дальнейшем мы будет употреблять его, но понимать в смысле биогеоценоза. Существование всех экологических систем обусловлено постоянным потоком энергии и круговоротом вещества в них.
Все организмы на основании своей функциональной роли в экосистемеподразделяются на три важнейшие группы – продуценты, консументы и редуценты.
Продуценты, или автотрофные организмы, создают органические вещества из неорганических в процессах фотосинтеза и хемосинтеза. Часть электромагнитной энергии солнечного излучения при этом переходит в энергию химических связей синтезируемых органических веществ.
Консументы, или гетеротрофные организмы, не способны создавать самостоятельно органические вещества из неорганических, поэтому они используют для своей жизнедеятельности органические вещества, созданные автотрофными организмами.
Редуцентами являются преимущественно одноклеточные гетеротрофные организмы (бактерии, протисты), а также некоторые многоклеточные (грибы, почвенные черви и др.), которые разлагают мельчайшие органические остатки до простейших неорганических соединений – H2О, CO2, N2 и др., вновь возвращая их в экосистему. Выделяющийся при фотосинтезе кислород используется для дыхания самих растений, а также гетеротрофных организмов. Тепловая энергия, выделяющаяся при дыхании автотрофных и гетеротрофных организмов,
рассеивается в пространстве, повышая энтропию окружающей среды. Образующиеся в процессах жизнедеятельности гетеротрофных организмов углекислый газ и вода используются автотрофными организмами в процессе фотосинтеза.
Впервые целостные представления о закономерностях трансформации вещества энергии в экологических системах разработал шведский ученый Р. Линдеман, который в 1942 г. опубликовал работу о превращениях солнечной энергии в одном из арктических озер Швеции. Тем самым были заложены основы нового направления экологии – экологической биоэнергетики, которая оказала значительное воздействие на прогресс экологии и биологии в целом. В дальнейшем это направление интенсивно разрабатывалось многими учеными во
всем мире. Среди них необходимо особо отметить Г. Г. Винберга, который в 1950–1960 гг. работал в Беларуси и стал основоположником белорусской экологической школы.
Продуценты, или автотрофные организмы, являются основой существования всех экологических систем и биосферы Земли в целом, поскольку только они способны создавать органические вещества из неорганических впроцессах фотосинтеза и хемосинтеза. По этому признаку продуценты подразделяются на фото- и хемоавтотрофов.
Трофическая структура экосистем. Процессы трансформации вещества и энергии в экосистеме происходят преимущественно посредством трофических связей (от греческого трофос – пища), т. е. в результате поедания одних организмов другими. Совокупность трофических связей в экосистеме образует трофические, или пищевые, цепи. Совокупность организмов, занимающих одинаковое положение в трофических цепях относительно их начала, называется трофическим
уровнем. Обычно каждый организм имеет несколько источников питания, находящихся на разных трофических уровнях, и сам может использоваться как объект питания организмами с различных уровней. Поэтому цепи питания в большинстве экосистем многократно разветвляются и образуют трофические сети. Различают пастбищные, детритные и паразитические пищевые цепи. Начальным звеном пастбищных цепей являются живые автотрофные
(фотосинтезирующие или хемосинтезирующие) организмы. В пастбищных пищевых цепях первый трофический уровень образуют продуценты – автотрофные организмы. Последующие трофические уровни образуют гетеротрофные организмы. Второй трофический уровень составляют растительноядные организмы (консументы первого порядка) – животные, гетеротрофные протисты и бактерии. Третий и последующие трофические уровни (консументы второго, третьего и т.д. порядка) образуют хищные организмы – почти исключительно животные. Обычно разные трофические уровни в экосистемах не разделены в
пространстве. Однако в некоторых случаях они достаточно четко дифференцированы. Например, это имеет место в геотермальных источниках суши, в которых температура воды у изливов горячих подземных вод достигает 50–60 оС и более. Автотрофные организмы, преимущественно цианобактерии, образуют здесь специфические перифитонные сообщества («маты»), которые распространены при температурах свыше 40–45 оС, поскольку при более низких температурах они не выживают. С другой стороны, гетеротрофные организмы (моллюски, членистоногие и др.) в геотермальных источниках не встречаются при температурах более 33–36 оС, поэтому они питаются фрагментами мата, сносимого течением воды в зоны с более низкой температурой. Отсюда в таких геотермальных источниках четко выделяется автотрофная зона, где распространены только автотрофные организмы, и гетеротрофная зона,
где автотрофные организмы отсутствуют и встречаются только гетеротрофные организмы (Хмелева, Голубев, Лаенко, 1984). В океанах и морях автотрофные организмы росли) существуют только до глубины проникновения света (максимум до 150–200 м). Гетеротрофные
организмы, обитающие в более глубоких слоях воды, ночью поднимаются к поверхности, чтобы питаться водорослями, а утром вновь уходят на глубину, совершая суточные вертикальные миграции протяженностью до 500–1000 м. В свою очередь, с наступлением утра гетеротрофные организмы из еще более глубоких слоев поднимаются наверх, чтобы питаться организмами, опускающимися из вышележащих слоев. Таким образом, в океанах и глубоких морях существует своеобразная «пищевая лестница», благодаря которой органическое вещество, создаваемое автотрофными организмами в поверхностных слоях воды, переносится по
цепочке живых организмов до самого дна. Вторым важнейшим источником органических веществ для видов из глубоководных экосистем являются организмы, отмирающие в верхних слоях и опускающиеся на дно («дождь трупов»). Общая масса отмирающих организмов в Мировом океане за год достигает не менее нескольких сотен миллионов тонн. В этой связи некоторые экологи рассматривают всю водную толщу отдельных регионов морей и океанов
как единый биогеоценоз. Другие ученые полагают, что условия среды в поверхностных и придонных слоях воды настолько различны, что они являются разными биогеоценозами.
В детритных пищевых цепях, или цепях разложения, продуценты (т. е. автотрофные организмы) отсутствуют. Их началом является детрит, или отмершее органическое вещество, представляющее собой остатки отмерших живых организмов. Детрит образует первый трофический уровень детритной пищевой цепи. Пастбищные пищевые цепи распространены преимущественно с сухопутных и морских экосистемах. Детритные цепи преобладают в
континентальных водоемах. Значительная часть пищевых ресурсов в них создается не в результате фотосинтеза (автохтонное органическое вещество), а поступает туда из наземных экосистем (аллохтонное органическое вещество), например, в виде листового опада. Гетеротрофные организмы, питающиеся непосредственно детритом, называются детритофагами. В наземных экосистемах ими являются многие виды насекомых, червей и др. Крупные детритофаги, к которым относятся некоторые виды птиц (грифы, вороны и т. д.), млекопитающих (гиены и прочие), а также многочисленные виды насекомых называют падальщиками. В водных экосистемах наиболее распространенными детритофагами являются
членистоногие – водные насекомые и их личинки и ракообразные. Детритофагами питаются другие, более крупные гетеротрофные организмы, которые, в свою очередь, могут служить пищей для хищников. Детритные пищевые цепи имеют огромное значение в природе. Если бы
их не было, то вся поверхность Земли быстро бы покрылась многокилометровым слоем отмерших организмов. Большое значение в разложении отмерших остатков растительного происхождения, особенно древесины, имеют грибы. Мельчайшие частицы отмершего органического вещества, которые не в состоянии усвоить детритофаги, разлагают бактерии и протисты, обладающие
благодаря своим мельчайшим размерам очень высокой интенсивностью метаболизма. По этой причине они разлагают органические остатки до простых неорганических соединений (углекислый газ, вода, аммиак, азот и др.). Эти вещества могут возвращаться в экосистему и вновь принимать участие в процессах круговорота веществ. Как и в пастбищных цепях, последний трофических уровень детритных пищевых цепей образуют редуценты-
микроконсументы. Практически любой вид живых организмов имеет своих паразитов. Начальным звеном паразитических пищевых цепей является свободноживущий организм-хозяин. На нем паразитируют паразиты первого порядка, а на них в, свою очередь, паразиты второго порядка и т. д. Аскариды, паразитирующие на пойкилотермных организмах, являются
паразитами первого порядка, паразитирующие в клетках аскарид одноклеточные эукариоты –паразитами второго порядка, обитающие в них бактерии – паразитами третьего порядка, паразитирующие в бактериях вирусы (бактериофаги) – четвертого порядка и т. д. В экологических системах все перечисленные типы пищевых цепей не существуют изолированно, а тесно взаимосвязаны. Так, отмирающие организмы из пастбищных пищевых цепей и неусвоенные остатки пищи живых организмов поступают в детритные цепи, поэтому в экосистемах формируется Y-образная
50.Любая совокупность организмов, существующих в
определенном объеме пространства, и их связей между собой и окружающей средой определяется как экологическая система, или экосистема. Термин «экосистема» впервые предложил английский эколог А. Тэнсли в 1935 г., хотя представления об экосистемах как особой форме взаимоотношений организмов со средой своего обитания возникли значительно раньше.Легко видеть, что понятие экосистемы является очень широким и не имеет ранга размерности. Экосистемой можно назвать любой объем пространства, в котором существуют живые организмы, хотя бы один. Ей может быть и капля воды с одной амебой, пруд, лесной массив, пилотируемый космический корабль и вся Биосфера Земли.
В 1944 г. русский геоботаник В. Н. Сукачев предложил более конкретный
термин «биогеоценоз». Он определяется как участок земной поверхности (в
том числе и водной) с определенной совокупностью живых организмов и
вполне однородными условиями среды, достаточно отделенный от других
участков, в пределах которого осуществляется более-менее замкнутый процесс круговорота веществ. Примерами биогеоценозов являются пруд, луг, лесной массив, болото и т. д.Термины «экосистема» и «биогеоценоз» достаточно близки, но не полностью тождественны, поскольку понятие биогеоценоза значительно более конкретно, чем экосистемы. Например, капля воды и Биосфера являются экосистемами, но не биогеоценозами. Однако поскольку термин «экосистема» легче произносить, в дальнейшем мы будет употреблять его, но понимать в смысле биогеоценоза.
Существование всех экологических систем обусловлено постоянным
потоком энергии и круговоротом вещества в них.
Все организмы на основании своей функциональной роли в экосистеме
подразделяются на три важнейшие группы – продуценты, консументы и
редуценты.Продуценты, или автотрофные организмы, создают органические вещества из неорганических в процессах фотосинтеза и хемосинтеза. Часть
электромагнитной энергии солнечного излучения при этом переходит в энергию химических связей синтезируемых органических веществ.
Консументы, или гетеротрофные организмы, не способны создавать
самостоятельно органические вещества из неорганических, поэтому они
используют для своей жизнедеятельности органические вещества, созданные
автотрофными организмами.
Редуцентами являются преимущественно одноклеточные гетеротрофные
организмы (бактерии, протисты), а также некоторые многоклеточные (грибы,
почвенные черви и др.), которые разлагают мельчайшие органические остатки до простейших неорганических соединений – H2О, CO2, N2 и др., вновь возвращая их в экосистему.
51. Различные типы объединений живых организмов. Живые организмы в
экосистеме группируются не хаотически, а объединяются в определенные
группы по причине своих биологических особенностей и воздействия
определенных факторов внешней среды. Выделяют несколько типов подобных объединений, отличающихся разной степенью интегрированности.
1. Общественные группировки, объединяющие особей одного вида. В
их образовании важнейшая роль принадлежит не факторам внешней среды, а
социальным отношениям и поведенческим реакциям. Они образуются у
насекомых, птиц, млекопитающих. Их примерами являются пчелиный рой,
муравейник, косяк рыб, прайд львов, сообщество обезьян и т. п.
Общественные группировки относительно независимы от окружающей
среды, они сохраняются даже тогда, когда условия среды меняются. Например,муравейник может сохраняться на протяжении нескольких десятков лет.
2. Скопления – собрание особей, относящихся к одному или
нескольким видам. Они возникают исключительно под воздействием одного или нескольких внешних факторов, выступающих в качестве центра притяжения.Скопления имеют случайный и временный характер. Между организмами,образующими скопление, нет устойчивых связей; оно разрушается, как только исчезает центр притяжения. Примером скопления может быть ночной рой насекомых, образованный особями разных видов, вокруг электрической лампы.Если лампу выключить, рой исчезает.
3. Активные паразитарные сообщества возникают благодаря
привлечению одного животного другим; оно происходит между разными
видами, но имеет исключительно односторонний характер. Например, любой
живой организм имеет многочисленных паразитов и комменсалов, которые
могут принадлежать к нескольким десяткам (если не сотням) видов. При гибели хозяина эти сообщества, как правило, распадаются.
4. Псевдообщественные сообщества – скопления, возникшие под
действием тигмотаксисов, т. е. стремления организмов к контакту с
определенным субстратом. Например, в водоемах одни группы организмов
обитают только на твердых, а другие – на мягких грунтах.
5. Биоценоз. Этот термин предложил немецкий зоолог Карл Мёбиус в 1877
г. в работе о сообществах животных и растений, приуроченных к устричным
банкам. Сам Мёбиус дал следующее пространное определение биоценозу –
объединение живых организмов, соответствующее по своему составу, числу
видов и особей некоторым средним условиям среды; объединение, в котором
организмы связаны взаимной зависимостью и сохраняются благодаря
постоянному размножению в определенных местах.… Если бы одно из условий отклонилось на некоторое время от обычной средней величины, изменился бы весь биоценоз …. Биоценоз также претерпел бы изменения, если бы число особей данного вида увеличилось или уменьшилось благодаря деятельности человека, или же один вид полностью исчез из сообщества, или, наконец, в его состав вошел новый».
Синонимами термина «биоценоз» являются такие понятия, как
«ассоциация», «сообщество» и «биотическое сообщество». Часто под
биоценозом понимается не совокупность организмов, а совокупность популяций.Термин «сообщество» можно также применить для обозначения любой группы взаимодействующих популяций. Например, можно рассматривать сообщества лужи, дубового листа и т. д. Однако мы будет понимать это пространство в смысле биоценоза.Совокупность популяций отдельных видов, населяющих биоценоз, носит название биоценотическое сообщество, или биоценоз. Природная среда, в которой существует биоценоз, называется биотоп. Поэтому биогеоценоз =биоценоз + биотоп.
Виды, образующие биоценоз, взаимно связаны друг с другом. Это отличает биоценоз от скоплений и псевдообщественных сообществ, где подобных связей нет или они очень слабы. Взаимосвязь эта так сильна, что
изменения, касающиеся только одного вида, могут отразиться на всем биоценозе и даже вызвать его распад. Особенно это касается видов-эдификаторов, или «строителей сообщества», которые в результате своей жизнедеятельности создают специфическую среду обитания для других видов сообщества. Примером вида-эдификатора может быть ель в сообществе старого и густого елового леса. Кроны еловых деревьев образуют сомкнутый полог, под которым создается специфический микроклимат, характеризующийся резким снижением освещенности, особым температурным режимом и повышенной влажностью воздуха. В таких условиях могут существовать только тенелюбивые и влаголюбивые виды живых организмов.
Биоценозы подразделяются на несколько уровней.
1. Основные сообщества. В Биосфере Земли их всего три – наземные,
пресноводные и морские.
2. Биомы, или формации, природные комплексы, природные зоны.
Биом – это однородное сообщество, не зависящее от состава растительности и занимающее обширную территорию – от миллионов до десятков миллионов км2.Причинами, обусловливающими видовой состав биома являются особенности климата, геологические факторы (преграды и пр.), эволюция живых организмов.Примерами биомов являются тундра, тайга, саванна, коралловые рифы,открытый океан, шельфовые зоны и т. п. Биомы в равной степени являются объектами изучения экологии и биогеографии.
3. Ассоциации, выделяемые в пределах биома, представляют собой
локальные, хорошо очерченные группировки видов. Их примерами являются
различные типы леса – хвойный, дубовый и т. п.
4. Синузии, или микроассоциации – некоторые очень небольшие
сообщества, четко ограниченные в пространстве и сохраняющие свою
индивидуальность. Ими могут быть виды, поселяющиеся на трупе крупного
животного, стволе мертвого дерева и т.п. Отдельные синузии существуют очень непродолжительное время, однако синузии, постоянно возникающие в пределах одной ассоциации, имеют сходный видовой состав.
10.6. Таксономический состав экологических систем. Все экологические системы существуют в пространстве и времени благодаря происходящим в них процессам трансформации вещества и энергии. В этих процессах принимают участие все имеющиеся в экосистемах живые организмы.
Все они значительно различаются по строению и особенностям метаболизма, в результате чего выполняют разные функции в экосистеме.На основе ряда фундаментальных особенностей строения и метаболизма все виды живых организмов на Земле подразделяются на ряд высших таксономических групп, которые имеют ранг доменов (domain), или надцарств и царств. Прежние достаточно простые схемы, выделяющие четыре или пять царств, в настоящее время кардинально пересмотрены. Сейчас все живые организмы подразделяются на три домена – Архебактери(Archaea), Бактерии
(Bacteria) и Эукариоты (Eucaria). Каждый домен включает не менее пяти групп,ранг которых соответствует царству.
1. Архебактерии (несколько сотен видов) – аэробные и анаэробные
прокариотные организмы. Обитают в глубовоководных районах Мирового
океана близи выходов гидротермальных вод. Отдельные виды обитают при
температуре свыше 114 оС. Способны к хемосинтезу.
2. Бактерии (около 80 тысяч видов) – прокариотные организмы, в клетках
которых отсутствуют ядра, митохондрии и хлоропласты. К ним относятся ряд групп бактерий и цианобактерии. Большинство – гетеротрофы с сапрофитным способом питания. Многие виды способны к фотосинтезу или хемосинтезу. Не менее 2–3 тысяч видов являются облигатными анаэробами.
3. Эукариоты (более 2 миллионов видов) – одноклеточные и
многоклеточные организмы, в клетках которых имеются ядра и митохондрии, а у автотрофных форм – и хлоропласты. К ним относится ряд царств одноклеточных, которых ранее объединяли в царство Протистов – Жгутиковые,Ресничные и Микроспоридии, а также «коронные» царства, включающие многоклеточные организмы, – Грибы, Растения и Животные. Все животные, грибы и многие протисты являются гетеротрофами. Способы их питания различаются от сапрофитного – у грибов и животных-эндопаразитов до голозойного (т. е. питания оформленной пищей) – у свободноживущих и протистов. Абсолютное большинство растений и многие протисты являются автотрофами.
Топические группировка биогеоценозов. В наземных экосистемах
растительные сообщества автотрофных организмов обычно представлены
сосудистыми растениями – покрытосемянными, голосемянными,
папоротникообразные, хвощами, плаунами, мохообразными, лишайниками. Эти сообщества обычно подразделяются на пять вертикальных ярусов. Первый, самый верхний, ярус составляют высокие деревья, кроны которых часто смыкаются, образуя сплошной полог. Второй ярус образуют кустарники(крушина, бузина, малина и др.), третий – кустарнички (черника, брусника,голубика, клюква, вереск и пр.). Четвертый ярус формируют травянистые растения и, наконец, пятый ярус – напочвенные мхи и лишайники.В некоторых сообществах отдельные ярусы могут отсутствовать.
Например, в старых и густых еловых лесах часто имеется лишь первый ярус.
Сомкнутые кроны елей создают хорошее затенение, что затрудняет развитие не только кустарниковой растительности, но и собственного подроста, а мощный слой хвойного опада препятствует формированию остальных ярусов.В высоких и густых тропических лесах микроклимат отдельных ярусов может существенно различаться. В приземном слое здесь очень высокая влажность и низкая освещенность, а температура в течение суток практически не изменяется. Напротив, микроклимат у верхушек деревьев, характеризующийся очень высокой освещенностью, более низкой влажностью и значительными суточными колебаниями температуры, больше напоминает климат саванны.В каждом из ярусов можно найти своеобразные группировки гетеротрофных организмов, преимущественно насекомых, но иногда к ним приурочены определенные виды птиц, млекопитающих (например, белки,ленивцы) и даже амфибий (квакши) и рептилий (летучий дракон).
В водных экосистемах существуют специфические биоценозы донных
организмов и обитающих в водной толще, называемые соответственно бентос и планктон. Далее сообщество донных бактерий называется бактериобентос,растений – фитобентос и животных – зообентос. По тому же принципу выделяют бактериопланктон, фитопланктон и зоопланктон.В морских экосистемах фитобентос образуют исключительно
многоклеточные водоросли – красные, бурые и др.; виды цветковых растений,например, зостера Zostera marina, здесь единичны. В фитобентосе пресных водоемов, напротив, доминируют погруженные и полупогруженные цветковые растения, а многоклеточные водоросли (харовые, зеленые и др.) имеют второстепенное значение.
Фитопланктон в обоих случаях представлен одноклеточными
водорослями; в морских экосистемах доминируют диатомовые, а в
пресноводных – зеленые водоросли. К пресноводному фитопланктону (а не к
бактериопланктону) также обычно относят и цианобактерий. Последние
периодически, обычно во второй половине лета, могут достигать очень высокой численности и биомассы («цветение воды») и тогда вытесняют остальные виды водорослей.
К зоопланктону относят достаточно мелких животных, не способных
противостоять течению воды, хотя многие их виды в стоячей воде способны к активному плаванию. В зоопланктоне пресных водоемов представлены
преимущественно мелкие формы – веслоногие и ветвистоусые ракообразные и коловратки. Зоопланктон морских водоемов значительно более разнообразен. В нем помимо мелких ракообразных (преимущественно веслоногих, мизид и эуфаузиид), аппендикулярий и других, присутствуют и значительно более крупные медузы и сальпы; последние, наряду с аппендикуляриями, относятся к разным подтипам хордовых. Значительную долю в зоопланктоне континентальных и морских водоемов составляют личинки многочисленных видов бентосных животных, выполняющие функцию расселения, а также личинки и мальки рыб.
Зообентос континентальных водоемов в видовом отношении достаточно
однообразен – в нем доминируют брюхоногие и двустворчатые моллюски,
членистоногие (почти исключительно ракообразные и насекомые и их личинки)
и малощетинковые черви. Напротив, в зообентосе морских водоемов
представлено большинство таксонов водных животных, поэтому его можно
здесь охарактеризовать лишь в самых общих чертах. В первую очередь
необходимо отметить коралловых полипов, которые как виды-эдификаторы в
экосистемах коралловых рифов создают среду обитания для многочисленных
видов морских водорослей, беспозвоночных и рыб. Значительная роль в
морском зообентосе также принадлежит двустворчатым и брюхоногим
моллюскам, кольчатым червям, ракообразным, иглокожим, асцидиям. В
зообентосе глубоководных океанических экосистем доминируют погонофоры.Сообщество водных организмов, постоянно обитающих в толще воды, но способных активно плавать и противостоять течению, называется нектон.Нектон пресных водоемов образуют почти исключительно рыбы. В нектонеморей и океанов наряду с ними присутствуют головоногие моллюски (кальмары,осьминоги) и морские млекопитающие (киты, дельфины, дюгони).
Сообщества, образующие обрастания на вертикальных естественных
(скалы, камни, подводные растения) и искусственных (сваи, подводные
сооружения, днища судов) подводных субстратах, называются перифитоном.Он подразделяется на зоо- и фитоперифитон.
Сообщества зоо- и фитоперифитона континентальных и морских
водоемов в таксономическом отношении достаточно сходны. В зооперифитоне присутствуют губки, мшанки, моллюски, нематоды, полихеты и др. Для морских сообществ характерно также наличие усоногих ракообразных и иглокожих. Видами-эдификаторами в разных сообществах зооперифитона могут являться двустворчатые моллюски, образующие устричные и мидиевые банки,губки, мшанки и др. Фитоперифитон образуют различные группы многоклеточных водорослей.Сообщества перифитона могут приносить значительный вред человеку.В них имеется ряд видов животных, которые разрушают не только подводные деревянные сооружении (например, двустворчатый моллюск – корабельный червь), но даже металлические конструкции – некоторые виды морских ежей.Обрастания на днищах кораблей значительно снижают их скорость.
52. Свойство экосистемы создавать в процессе своего
функционирования органическое вещество определяется как биологическая
продуктивность. Органическое вещество, создаваемое в экосистеме, в
результате прироста массы тела населяющих ее организмов, называется
продукцией. Различают продукцию популяций отдельных видов, видовых
сообществ, целых трофических уровней и всей экосистемы.
Все органическое вещество, создаваемое автотрофными организмами в
процессах фотосинтеза или хемосинтеза, называется валовой первичной
продукцией (ВПП). Однако значительная часть синтезированного
органического вещества сразу же расходуется ими в процессах метаболизма.
Разность между энергией ВПП и расходом ее в метаболических процессах (М)является чистой первичной продукцией (ЧПП). Таким образом,
ЧПП = ВПП – М.Чистая первичная продукция является тем самым органическим веществом, которое накапливается в растениях. Из него в результате сложных процессов, происходящих в растениях, образуются все их органы – стебли, листья, корни, цветки плоды и другие, которые могут использоваться в пищу гетеротрофными организмами.
Органическое вещество, создаваемое гетеротрофными организмами,
находящимися на втором и последующих трофических уровнях, называется
вторичной продукцией. Она равна сумме приростов массы их тела
гетеротрофных организмов за определенный период времени. В большинстве
случаев вторичная продукция, которую часто называют просто продукцией,
рассчитывается для популяций отдельных видов. Этим она отличается от
первичной продукции, которую возможно рассчитать для всего сообщества
автотрофных организмов, составляющих данную экосистему.
Количество органического вещества, покидающего тем или иным
способом экосистему (сбор урожая, вылов рыбы, вылет наземных насекомых
из их водных личинок и т. д.), называется конечной продукцией.
Способы выражения биомассы и продукции. Общее количество живого
вещества в экосистеме называется биомассой. Биомасса экосистемы
образована всеми населяющими ее живыми организмами. Продукция
экосистемы равна приросту ее биомассы за определенный промежуток
времени. Биомасса и продукция экосистем могут быть выражены в различных единицах, однако предпочтительнее их выражать в одинаковых единицах.Биомасса экосистемы рассчитывается как произведение численности (или плотности) особей в экосистеме на их среднюю массу. Поскольку особи в экосистеме объединены в популяции отдельных видов, биомасса экосистемы равна сумме биомасс всех составляющих ее популяций. В свою очередь биомасса популяции равна сумме масс тела всех ее особей. Биомасса экосистемы является статическим показателем, который не имеет размерности времени. Напротив, продукция – динамический показатель,который определяется для какого-либо промежутка времени – суток, месяца,года и т. д. В наземных экосистемах биомасса и продукция обычно пересчитываются на единицу площади (г·м-2; т·км-2·год-1 и т. д.).
Одним из важнейших показателей, характеризующих продукционные
возможности организмов, является Р/B-коэффициент, или отношение
продукции (Р) какой-либо популяции или даже всего трофического уровня к
биомассе (В) этой популяции или трофического уровня. Значения Р/B
рассчитываются для определенного периода времени, обычно за сутки, месяц,сезон вегетации, год и т.д., поэтому имеют размерность соответственно год-1,сезон вегетации-1, месяц-1, сутки-1 и т. д.
Суточный Р/B-коэффициент популяций гетеротрофных организмов часто
называют удельной продукцией. В качестве биомассы при расчетах Р/B
используют ее минимальное или, максимальное, но чаще среднее значение за
тот период времени, для которого производится расчет.
53. Различают валовую П. п., равную общему кол-ву продуктов фотосинтеза за определённый отрезок времени, и чистую П. п., равную разности между валовой П. п. и той её частью, к-рая была затрачена в процессе дыхания. Все органическое вещество, создаваемое автотрофными организмами в
процессах фотосинтеза или хемосинтеза, называется валовой первичной
продукцией (ВПП). Методы определения первичной продукции. Чистая первичная продукция наземных экосистем обычно оценивают по годовому приросту биомассы растений. Чтобы определить ее величину за сезон вегетации, в конце этого сезона все растения, находящиеся на определенных территориях известной площади, удаляют и взвешивают. Поскольку обычно корни и подземные части растений не учитываются, определяют только массу их наземных органов – или чистую годовую наземную продуктивность. По отношению зерновым сельскохозяйственным культурам она будет равной суммарной массе собранного зерна, соломы, ботвы и т.п.
В балансовом уравнении фотосинтеза соотношение молекулярной массы
одной молекулы глюкозы и шести молекул кислорода составляет 180:192 =
0,9375.
6СО2 + 6Н20 → С6Н1206 + 02 .
264 108 180 192
Поэтому по величине прироста органической массы растений (без учета
воды и минеральных веществ) за определенный период времени возможно
приблизительно оценить массу произведенного ими кислорода
Пример. Общая урожайность кукурузы (зеленая масса, корни, початки и
пр.) достигает 500 ц·га-1. Содержание воды в кукурузе равно в среднем 50%. Отсюда за период выращивания кукурузы на одном гектаре произведено (500 · 0,5)/0,9375 = 267,5 ц кислорода.
Первичную продукцию в водных экосистемах определяют методом
«темных склянок». Суть его состоит в следующем. Вода из водоема
пропускается через тонкий фильтр, чтобы удалить из нее зоопланктон, затем в ней определяют исходное содержание кислорода. После этого вода заливается в одинаковые бутылочки из светлого темного стекла (не пропускающего солнечные лучи), которые опускаются в водоем на сутки или другие промежутки времени. В светлых склянках происходят процессы фотосинтеза и дыхания фитопланктона, в темных – только его дыхание. Отсюда ЧПП фитопланктона равна разности между содержанием кислорода в светлой в склянке и контроле, а его дыхание – разности между содержанием кислорода в контроле и темной склянке.
Пример. Начальное содержание кислорода в воде (контроль) равно 6 мг·л-1, в
конце опыта в светлой склянке оно возросло до 8 мг·л-1, а в темной понизилось до 3
мг·л-1. Отсюда ЧПП фитопланктона составит 8 – 6 = 2 мгО2·л-1, а его дыхание - 6
– 3 = 3 мгО2·л-1. Тогда ВПП фитопланктона (ЧПП + Т) равна 2 + 3 = 5 мгО2·л-1.
Радиоуглеродный метод определения первичной продукции основан на
том, что живые организмы в равных пропорциях поглощают из среды обычный нерадиоактивный изотоп углерода (12C) и его радиоактивный изотоп (14С) с периодом полураспада 5600 лет.В какую-либо герметичную установку добавляют известное количество изотопа 14С, например, в виде 14СО2. Затем после определенной экспозиции определяют содержание 14С в растении. Чистую первичную продукцию рассчитывают по формуле
ЧПП = A r/(Cr/Сs).
где Cr – содержание 14CO2 в начале эксперимента, Сs – общее содержание
CO2 в начале эксперимента (14CO2 + 12CO2), Ar – количество 14CO2,
поглощенного растениями.
Пример. В установку внесли 100 мг смеси (14CO2 + 12CO2), в которой
соотношение Cr/Сs = 0,10, т. е. количество 14CO2 составило 10 мг.
Величина потребления 14CO2 за сутки составила 2 мг. Тогда ЧПП можно
рассчитать из пропорции: 10 : 2 = 100 : Х. Отсюда Х = (100 . 2)/10 = 20.
Еще одним методом определения первичной продукции в водных
экосистемах является ее расчет по содержанию хлорофилла а в планктоне.
Для этого фитопланктон из известного объема воды концентрируют
фильтрованием на мембранном фильтре, фильтрат высушивают, а затем из него экстрагируют фотосинтетические пигменты в определенном объеме 90%-ного ацетона. Хлорофилл а характеризуется определенным спектром поглощения,поэтому его концентрацию в экстракте можно определить на спектрофотометре по величине экстинкции (ослаблению) света (Бульон, 1981). Рабочей группой экспертов ЮНЕСКО предложено следующее уравнение для расчета концентрации хлорофилла а (Схл, мг·м-3) в воде:
Схл = (11,64Е663 – 2,16 Е645 + 0,1 Е630) · (v/VL),
где Е663 , Е645 и Е630 – экстинкция света при длине волн соответственно
663, 645 и 630 нм; v – объем экстракта (мл); V – объем пробы воды (л); L –
длина пробега светового луча в экстракте (см).
Пример. В водоеме взята проба воды объемом (V) 0,5 л. Фотосинтетические
пигменты экстрагированы в 5 мл ацетона (v). На спектрофотометре в кювете длиной 1см (L) получены следующие значения экстинкции экстракта: Е663 = 0,048; Е645 =0,028; Е630 = 0,008 (по Алимову, 1989). Отсюда концентрация хлорофилла а в исследованной пробе воды равна
Схл = (11,64 · 0,048 – 2,16 · 0,028 + 0,1 · 0,008) · (5/0,5 ·1) = 5,28 мг м-3.
Поскольку молекулы хлорофилла присутствуют только в живых клетках,
рассчитанные значения Схл являются также хорошим показателем биомассы
сообществ фитопланктона. Для расчета первичной продукции по содержанию хлорофилла необходимо знать ассимиляционное число (АЧ), представляющее отношение массы органического вещества, синтезированного в единицу времени, к массе хлорофилла а в сообществе фитопланктона.АЧ принято выражать в мг углерода (С) на 1 мг хлорофилла а за час или за сутки. Максимальные значения АЧ для озер разных широт изменяются в достаточно узких пределах – от 1 до 10 мгС/мг хлорофилла а в час, а его среднее значение на глубинах с оптимальными для фотосинтеза условиями среды можно принять равным 2 мгС/мг хлорофилла а за час (Алимов, 1989).Первичная продукция для этого слоя воды определяется умножением концентрации хлорофилла а в нем на АЧ.
54. Фотосинтез — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов.Сущность синтеза органических веществ состоит в переносе атомов водорода и углерода на атомы углерода какого-либо другого соединения, в
результате чего происходит удлинение его углеродных цепочек. Поскольку эти реакции идут с затратой энергии, в процессе фотосинтеза для их протекания используется энергия солнечного светаРазличают оксигенный и неоксигенный фотосинтез. В первом случае донором водорода является вода и происходит выделение кислорода, во втором– донорами водорода являются другие вещества, а кислород не выделяется.Оксигенный фотосинтез имеет место у всех автотрофных эукариотов(протисты, низшие и высшие растения) и некоторых прокариотов(цианобактерии, или сине-зеленых водоросли). Неоксигенный фотосинтезвстречается только у немногих прокариотов – пурпурных и зеленых бактерий.
Оксигенный фотосинтез у эукариотов происходит в органеллах клетки –
хлоропластах, где содержатся особые пигменты (хлорофиллы (а, в, с и d) и
некоторые другие), улавливающие солнечный свет. У цианобактерий они
находятся во внемембранных органеллах, называемых хлоросомами,
соединенных с цитоплазматической мембраной.
Хлорофиллы, несколько различающиеся по строению и спектрам
поглощения (рис. 1), улавливают энергию видимого света в диапазоне длин
волн 380–710 нм. На этот диапазон, называемый физиологически активной
радиацией (ФАР), приходится около 45 % падающей на земную поверхность
энергии солнечной радиации; примерно столько же составляет инфракрасное,или тепловое излучение, а оставшиеся 10 % – ультрафиолетовое излучение
(Бигон и др., 1989).
Основным пигментом фотосинтеза является хлорофилл а, максимум
поглощения им света находится в пределах длин волн 600–700 нм. Хлорофиллы остальных типов являются дополнительными. Они не участвуют непосредственно в процессе фотосинтеза, но передают поглощенную ими световую энергию хлорофиллу а. Поскольку максимумы их поглощения отличаются от такового у хлорофилла а, это позволяет автотрофным организмам более эффективно использовать энергию всего диапазона ФАР. Молекула хлорофилла а поглощает квант света, который выбивает из нее один электрон. В таком состоянии она становится способным отобрать электрон от молекулы воды, в результате этого происходит ее фотолиз, т.е. распад на ион водорода (Н+) и гидроксильную группу (ОН-):
Н20 → H+ + OH-
и далее
4ОН- → 2Н20 + О2↑.
Важно отметить, что кислород, образовавшийся в процессе фотолиза воды,
является побочным продуктом фотосинтеза. Он не участвует в процессах
образования органического вещества, а выделяется в атмосферу. Тем не менее,роль кислорода в биосфере огромна, поскольку он используется для дыхания всеми аэробными организмами.Образующие в процессе фотолиза ионы водорода наряду с углекислым газом (СО2) участвуют в синтезе органических веществ. Суммарное уравнение фотосинтеза можно представить в виде
6СО2 + 12Н20 → С6Н1206 + 6H2O + 602↑ + 675 ккал.
Следовательно, на каждые 6 грамм-молекул СО2 и Н2О выделяется 6
грамм-молекул О2, а в образованной грамм-молекуле глюкозы (С6Н1206)
накапливается около 675 ккал энергии. При фотосинтезе образуется также
небольшое, в расчете на одну молекулу С6Н1206, количество молекул АТФ.
Часть органического вещества, образованного в процессе фотосинтеза,
сразу же расходуется растениями в процессе дыхания. Молекул АТФ,
образующихся непосредственно в процессе фотосинтеза, явно недостаточно для обеспечения протекания всех химических реакций в их клетках, тогда как в процессе аэробного дыхания при окислении одной молекулы С6Н1206
образуется до 38 молекул АТФ. Интенсивность дыхания растений днем и ночью приблизительно одинакова, однако фотосинтез происходит только в светлое время суток. Поэтому растения расходуют на дыхание в среднем до половины синтезируемых ими органических веществ.У растений существует и другой процесс – фотодыхание, или окисление органических соединений кислородом на свету до СО2 и Н2O без образования АТФ. Биологический смысл фотодыхания не вполне ясен. Предполагается, что оно может выполнять защитную функцию, связывая излишки кислорода, образующегося при фотосинтезе или окисления избытка восстановленных
органических соединений (Брода, 1978).
Типы оксигенного фотосинтеза выделяются по способу фиксации
растениями СО2. У большинства видов растений он идет по С3-пентозфосфатному пути, или циклу Кальвина. Однако у некоторых видов
фиксация СО2 происходит по С4-дикарбоновому пути. Поэтому эти группы
растений названы С3- и С4-растениями. Типичными С3-растениями являются
пшеница, рожь, фасоль, клевер, горошек, а С4-растениями – кукуруза, сахарный тростник, просо, сорго (Одум, 1986).
Особый тип фотосинтеза встречается у растений – обитателей жарких
пустынь, например, толстянковых, кактусов, молочаев и т. п. Их устьица в
светлое время суток закрыты, чтобы предотвратить потерю воды через
испарение, и открываются только прохладной ночью. Углекислый газ,
поглощаемый ночью через устьица, накапливается в клетке в форме
органических кислот и фиксируется в углеводах при наступлении светлого
времени суток. Такой тип фотосинтеза получил название САМ-метаболизм, от Crassulaceae acid metabolism, т. е. кислотный метаболизм растений семейства толстянковых.
Неоксигенный, или бактериальный фотосинтез имеет место у зеленых и пурпурных бактерий. Его пигментами являются бактериохлорофиллы a, b, c и d. Максимум их поглощения лежит в диапазоне длин волн от 800 до 900 нм, т. е. за пределами диапазона ФАР у эукариотов, в области невидимой для
человеческого глаза инфракрасной части спектра. Поэтому виды с оксигенным и неоксигенным фотосинтезом в экосистемах не конкурируют между собой за свет.При бактериальном фотосинтезе донором водорода является не вода, а другие вещества, как неорганические, например, сероводород (Н2S) и молекулярный водород (Н2), так и достаточно восстановленные органические соединения. Поэтому его побочными продуктами являются не свободный кислород, а элементарная сера (S) или ее соединения, например, серная кислота
(Н2SО4), вода или более окисленные органические соединение, например:
6Н2S + 12СО2 + 6Н2О→ 2С6Н1206 + 6Н2SO4
6СО2 + 12Н2 → С6Н1206 + 6Н2O.
6СО2 + 6СН3CHOHCH3 → С6Н1206 + 6CН3COCH3.
Большинство видов с бактериальных фотосинтезом – облигатные
анаэробы. Поэтому образование АТФ у них происходит при разложении
синтезированных органических соединений в процессах брожения.
Недавно у галофильных бактерий рода Halobacterium, обитающих в
пересоленных водоемах, был открыт принципиально новый способ образования АТФ. У них имеется пигмент бактериородопсин, сходный со зрительным пигментом сетчатки глаза – родопсином, который способен синтезировать АТФ,используя энергию поглощаемого им солнечного света.
Интенсивность фотосинтеза (ИФ) у растений в значительной степени зависят от факторов среды, в первую очередь от температуры и освещенности. При слабой и умеренной освещенности (обычно до 25 % от освещенности в яркий солнечный день) ИФ возрастает прямо пропорционально интенсивности освещения. Однако при более ярком свете рост ИФ начинает замедляться, затем стабилизируется на постоянном уровне, а потом резко снижается.
При постоянном освещении ИФ вначале повышается с увеличением
температуры, достигая определенного максимума в пределах сравнительно
узкого интервала температур. При дальнейшем повышении температуры ИФ
фотосинтеза резко падает. Температура, при которой отмечается максимальный фотосинтез, у растений различается от приблизительно 16 оС у видов из умеренных зон до 38 oC – у тропических видов.У С3-растений максимальная ИФ наблюдается при умеренной освещенности и температуре, высокие уровни освещенности и повышенная температура подавляют этот процесс. Напротив, С4-растения адаптированы к высокой температуре и яркому свету, а их фотосинтез не ингибируется высокими концентрациями кислорода. Кроме того, С4-растения меньше нуждаются в воде; как правило, на образование 1 г сухого вещества им требуется менее 400 г воды, а С3-растениям – от 400 до 1000 г воды. Поэтому в условиях жаркого и засушливого климата С4-растения значительно превосходят по продуктивности С3-растения. Однако последние получают преимущество при
слабой освещенности и низкой температуре.
У некоторых С3-растений на фотодыхание расходуется до 50 %
образующегося при фотосинтезе кислорода. Напротив, у С4-растений
фотодыхание не обнаружено, чем в значительной степени объясняется их
высокая урожайность, особенно таких культур, как кукуруза и сахарный
тростник.Очевидно, длительное существование фотосинтезирующих организмов возможно лишь, когда их величина их валовой первичной продукции (ВПП)превышает их метаболические траты на дыхание (T) или, по меньшей мере,равна им. Уровень освещенности, при котором ВПП = T, т. е. когда чистая первичная продукция растений равна нулю, называется световой компенсационной точкой. На суше световой фактор редко является
лимитирующим для распространения растений, поскольку освещенность в
светлое время суток даже в условиях затенения, как правило, выше световой
компенсационной точки. Напротив, в водоемах световой поток значительно ослабляется с глубиной и с увеличением содержания взвешенных органических веществ в воде. В морских водоемах с чистой водой наибольшей проникающей способностью обладают лучи зеленого и синего цветов. Поэтому наибольшей глубиной распространения в морях и океанах характеризуются красные водоросли, световая компенсационная точка для которых находится на глубинах до 100 – 200 м. Это объясняется тем, что помимо хлорофиллов, у красных водорослей имеются дополнительные пигменты фотосинтеза – красный фикоэритрин и синий фикоцианин, поглощающие световые волны в этих участках спектра.Еще одним важнейшим фактором, определяющим ИФ у растений, является индекс листовой поверхности (ИЛП), или отношение общей площади листьев у растений к площади соответствующего участка почвы, на котором эти
растения находятся. Значения ИЛП существенно различаются в разных
растительных сообществах
55. Хемосинтез. Большинство хемоавтотрофов является облигатными
аэробными бактериями, которые синтезируют органические вещества за счет
энергии, выделяющейся при кислородном окислении некоторых неорганических веществ. Хемоавтотрофы подразделяются на ряд основных групп соответственно природе окисляемых субстратов. У водородных бактерий процесс хемосинтеза можно выразить следующим
общим уравнением:
36Н2 + 12О2 + 6СО2 → С6Н12О6 + 30Н2О
Нитрифицирующие бактерии используют для синтеза органических
соединений окисление восстановленных соединений азота
NH4+ + 3/2O2 → NO-2 + H2O + 2H+ +
и далее
NO2- + ½O2 → NO-3
Данный процесс также часто называют нитратным дыханием.
Образующиеся при окислении NH4+ ионы водорода и выделяющаяся энергия
используются бактериями для фиксации СО2 в процессе, аналогичном темновой фазе фотосинтеза у С3-растений (цикл Кальвина).
Серобактерии в аэробных условиях окисляют соединения серы (например,
сероводород) , до серы:
2H2S + 02 → 2Н2O + 2S
и далее до серной кислоты:
2S + O2 + H20 → H2SO4
Выделяющаяся при этом энергия аккумулируется в молекулах АТФ.
Железобактерии окисляют двухвалентное сернокислое железо до
трехвалентного, согласно уравнению
4Fe2+ + 6SO4
2- + 4H+ + O2→ 2Fe2(SO4)3 + 2H2O
Эта реакция сопровождается очень малым выходом энергии, поэтому для
синтеза органических веществ железобактериям приходится окислять большое количество сернокислого железа – не менее 500 г на 1 г образованной бактериальной биомассы. Для сравнения, серобактерии для образования 1 г биомассы окисляют только 10 г серы.В последние годы открыты новые группы хемосинтезирующих организмов. Архебактерии гидротермальных источников. В 1980-х годах на дне
Атлантического и Тихого океанов на глубинах 2–3 км были обнаружены
скопления гидротермальных источников. Оттуда изливаются воды из верхнего слоя мантии Земли с температурой до 350 оС и высоким содержанием сероводорода, сероводорода и других соединений серы. Поскольку эти воды имеют темный цвет, гидротермальные источники получили образное название «черные курильщики».
Вокруг «черных курильщиков» сформировались уникальные экосистемы,
в которых обитает множество новых для науки видов бактерий и животных. В зонах с температурой воды до 115 оС существуют аэробные серобактерии. Они окисляют метан и сероводород, а выделяющуюся при этом энергию использует для образования АТФ с последующим синтезом органических веществ. Процесс их хемосинтеза можно схематично представить в следующем виде:
Н2S + O2 + CO2 → C6H12O6 + H2SO4
Литотрофные бактерии встречаются в породах известняка на глубинах
до 3 км. В среде, где они обитают, находятся лишь незначительные количества биогенных элементов. Предполагается, что они разлагают углекислый кальций(СаСО3 → СаО + СО2), а выделившуюся при этом энергию и углекислый газ используют для синтеза органических соединений.
Гидротрофные бактерии обитают в гранитных породах с повышенным
содержанием радиоактивных изотопов (235U, 238U и др.) до глубин 4–5 км.
Предполагается, что они используют для синтеза АТФ энергию, выделяющуюся при соединении ионов Н+ и ОН-, образующихся в процессе радиолиза воды в их цитоплазме γ-квантами, выделяющимися при распаде радиоактивных изотопов.
Фотосинтез и хемосинтез – единственные процессы в биологических
системах, в которых происходит запасание энергии.
56. Продукция популяций первого типа (у видов с коротким, до
нескольких месяцев, жизненным циклом и однократным размножением в
конце сезона вегетации) может быть рассчитана способами, аналогичными
приведенными для популяций второго типа. За весь период существования
популяции, обычно соответствующий длительности сезона вегетации, ее
продукция рассчитывается согласно уравнению (11.2). Поскольку биомасса
популяции в конце сезона вегетации (В2) равна нулю, его можно упростить:
Р = Ве – В1. (11.4)
Как правило, масса новорожденной молоди очень мала по сравнению с
массой взрослых особей. Поэтому начальная биомасса популяции (В1)
составляет незначительную часть от биомассы элиминированных особей (Ве),
следовательно Р » Ве. Таким образом, продукция популяций первого типа
приблизительно равна ее элиминации.
Допустим, снижение численности такой популяции (Nw) с увеличением
средней массы ее особей (W) соответствует экспоненциальному уравнению
(рис. 2):
Nw = Noe-aW, (5)
где No – начальная численность особей, а – эмпирический коэффициент. Приняв значение Wo = 0 и проинтегрировав (11.6), получаем значение
продукции когорты (Ве) за период времени (t) от появления новорожденных
особей до того, как их средняя масса достигла значения Wt:
Wo
Ве = (- No/a)e-aW ½| = (- No/a)(е-aW – 1) = (No/a)(1 – e-aWt).
0
Прибавив полученное значение Ве к биомассе выживших особей в
когорте, получим значение продукции популяции.
Пример. Функциональная связь между численностью когорты (N, экз·м-2) и
средней массой ее особей (W, мг), представленная на рис. 11.2, следует уравнению:
N = Noe-0,69W. Когда средняя масса особей в когорте достигла 100 мг, численность
когорты снизилась до 1 экз·м-2. Тогда биомасса элиминированных особей в когорте за
период времени от ее появления до того, до возраста, когда W = 100 мг, будет равна:
Ве = (1000/0,69)(1 – e-0,69·100) » 1000/0,69 » 1449 мг·м-2.
Биомасса когорты в конце периода ее роста равна 100 мг · 1 экз·м-2
= 100 мг·м-2. Тогда продукция когорты за указанный период времени составит
1449 мг·м-2 ·100 мг·м-2 = 1549 мг·м-2.
57.Продукция популяций второго типа (с жизненным циклом более
одного года и с однократным в году периодом размножения) в простейшем
случае при отсутствии смертности может быть рассчитана по формуле:
P = (W2 – W1)N = W2N– W1N = В2 – В1, (1)
где W1 – начальная масса особей, W2 – их конечная масса, N – численность
особей. Здесь предполагается, что в период времени, в котором средняя масса особей возрастает от W1 до W2, пополнения популяции молодью не
происходит.
Таким образом, при отсутствии смертности продукция популяции (Р) за
определенный период времени равна разности между конечной (В2) и
начальной (В2) ее биомассой. Однако поскольку в любых популяциях всегда
имеет место смертность, то полученная согласно (11.1) величина продукции
представляет собой максимально возможную, или потенциальную, продукцию популяции (Pmax).С учетом смертности расчет продукции несколько усложнится:Р = Ве + В2 – В1, (2)
где Ве – биомасса элиминированных особей, В2 и В1 – конечная и начальная
биомасса популяции. По (11.2) обычно рассчитывают продукцию популяций
долгоживущих видов за астрономический год.
Если величины смертности у особей разной массы или возраста
достаточно близки, то Ве можно рассчитать согласно:Ве = (N1 – N2) ½ [(B1/N1 + B2/N2)], (3)
где N1 и N2 – соответственно начальная и конечная численность популяции.
Легко видеть, что в (11.3) B1/N1 соответствует начальной массе особей
(W1), а B2/N2 – их конечной массе (W2). Отсюда выражение ½[(B1/N1 + B2/N2)]
соответствует средней массе элиминированных особей (We). Тогда
Ве = (N1 – N2)We.Приведенный способ расчета продукции часто называют также методом Бойсен-Иенсена, который впервые в 1911 г. рассчитал подобным образом продукцию популяций ряда видов двустворчатых моллюсков в двух бухтах Лим-фьорда (Дания).Расчет продукции популяции согласно (2) и (3) можно представить графически (рис.1).
Рис.
Легко видеть, что если число особей в популяции линейно снижается с
увеличением их средней массы (т. е. относительная смертность у особей всех
размеров постоянна), то значение Be, рассчитанное по (11.3), соответствует
площади треугольника N1N2N2
’. В свою очередь, площадь прямоугольника
N2W1W2N2
’ равна разности между конечной и начальной биомассой популяции,
т. е. В2 – В1.
Отсюда продукция популяции, рассчитанная согласно (11.2), равна
площади многоугольника N1W1W2N2’. Площадь большого прямоугольника
N1W1W2N1’ соответствует ее потенциальной продукции популяции (Pmax), т. е. продукции в отсутствии смертности.
Таким образом, продукция популяции второго типа соответствует
площади под кривой зависимости изменения численности особей в популяции
от их средней массы. Рассчитанное значение продукции относится к тому
промежутку времени, за который средняя масса особей в популяции изменится от W1 до W2.Однако в большинстве популяций величины смертности изменяются с массой и возрастом особей, обычно они выше у более молодых и меньших по размерам особей. Тогда изменение численности популяции с увеличением средней массы особей будет передаваться ниспадающей ломаной линией. В этом случае величины Ве определяют для более коротких интервалов массы особей, в которых смертность принимается постоянной. Затем величины Ве для отдельных интервалов суммируют, получая суммарное значение элиминации. Если в начале и конце расчетного периода времени биомасса популяции остается постоянной (В1 = В2), такая популяция называется стационарной.Поскольку в этом случае Р = Ве,, то продукция стационарной популяции
второго типа равна ее элиминации.
58. Продукция популяций третьего типа (у видов с постоянным
пополнением в течение сезона вегетации) рассчитывается следующим образом.
1. Через определенные промежутки времени из природной популяции
отбираются пробы особей для определения сезонной динамики ее размерной
структуры. Предполагается, что соотношение размерных групп особей в
отобранных пробах адекватно отражает это соотношение в природной
популяции. Пример сезонной динамики размерной структуры популяции
понтокаспийской мизиды Paramysis lacustris приведен ранее (рис.6).
2. Каждая отобранная проба разделяется на ряд размерных или весовых
групп, для каждой из которых определяется численность особей (Ni) и средняя
масса их тела (Wi).
3. Реконструируется усредненная кривая роста особей за весь жизненный
цикл. На ее основании определяются величины среднесуточного прироста
массы тела (Ci = ΔWi/Δt) или удельной скорости роста (Сwi = ΔWi/ΔtWi) у
особей с разной массой тела (Wi).
4. Рассчитывается функциональная связь между Ci или Сwi и Wi .
Суммарный суточный прирост массы тела в одной размерной группы
особей в популяции со средней массой Wi, равен:
Pi = Ni ΔWi /Δt. (7)
Умножив и разделив выражение Ni ΔWi /Δt в (7) на Wi, получаем:
Pi = Ni Wi ΔWi /Δt Wi. (8)
Поскольку в (11.8) значение NiWi соответствует биомассе особей i-той
размерной группы (Вi), а ΔWi/ΔtWi – удельной скорости роста особей данной
группы (Сwi), получаем равенство (11.9), эквивалентное (11.7):
Pi = ВiСwi. (9)
Среднесуточная продукция всей популяции (Р) равна сумме
среднесуточных приростов особей всех размерных групп, рассчитанных по (7)
или (8), т.е.:
Р = ΣPi.
Пример расчета продукции для одного из видов планктонных
ракообразных приведен в табл. 1.
Таблица 1. Расчет суточной продукции популяции веслоногого рака Cyclоps
colensis в озере Нарочь (по Методы…, 1968 c изменениями)
Весовая группа, мг Wi
|
Средняя масса, мг Wi |
Число особей, экз м-3 Ni
|
Средний прирост массы особи, мкг сутки-1 Сi
|
Биомасса, мкг м-3 Bi = WiNi
|
Продукция весовой группы, мкг м-3сутки-1 Pi = CiNi
|
||
0 – 5 |
2,5 |
750 |
0,07 |
1875 |
52,5
|
||
5 – 10 |
7,5 |
575 |
0,15 |
4313 |
86,2
|
||
10 – 25 |
17,5 |
625 |
0,29 |
10 938 |
181,2
|
||
25 – 40 |
32,5 |
130 |
0,87 |
4225 |
113,1
|
||
40 – 80 |
60,0 |
100 |
1,06 |
6000 |
106,0
|
||
Суммарные значения |
27 351 |
539,0
|
|||||
Из табл. 1 следует, что биомасса популяции Cyclops colensis на момент
взятия пробы равна 27 351 мкг·м-3, а ее продукция – 539 мкг·м-3·сутки-1; тогда
суточный Р/B-коэффициент популяции составляет 539 : 27 351 = 0,0197.
Продукцию популяции (ΣР) за периоды времени, превышающие одни
сутки (t2 – t1), определяют согласно
ΣР = ½ (Р2 + Р1)(t2 – t1),
где Р1 и Р2 – суточная продукция популяции в моменты времени t1 и t2 .
Затем значения ΣР для последовательных временных интервалов суммируют,
получая величину продукции для более продолжительных интервалов времени,
например, для всего сезона вегетации.
59. Продукции популяций четвертого типа. У ряда групп организмов масса тела в жизненном цикле изменяется незначительно. К ним относятся бактерии, свободноживущие протисты (амебы, инфузории и пр.), которые размножаются делением надвое, а также коловратки.Продукция одноклеточных организмов, размножающихся делением надвое, за единицу времени определяется следующим образом:
Р = rNW = rВ,
где r – удельная скорость роста численности популяции (время-1); N – средняя численность особей в популяции; W – средняя масса одной особи (Алимов,1989). Тогда произведение NW представляет биомассу популяции (B). Значение r рассчитывается согласно:r = ln2/g,
где g – время удвоения численности популяции, которую можно принять
равной среднему промежутку времени между двумя делениями одной особи.
Тогда продукция за единицу времени, соответствующую размерности r (час,
если промежуток между двумя делениями выражен в часах; сутки – если в
сутках и т.п.), равна:Р = (1/g) ln2•B.
Продукцию популяции за более длительные промежутки времени можно
рассчитать следующим образом. Примем, что средняя масса одной особи в
популяции постоянна. Тогда увеличение биомассы популяции во времени (t)
будет пропорционально росту ее численности.
Пусть начальная биомасса популяции (в момент времени t = 0) равна Во.
Поскольку одноклеточные организмы размножаются делением пополам,
биомасса их популяций (при отсутствии смертности) будет расти
экспоненциально. Тогда ко времени t биомасса популяции (Вt) составит:
Bt = Boe(ln2/g)t. (10)
Отсюда продукция популяции за период времени t (Рt) равна разности
между Вt и Bo, т. е.
Pt = Boe(ln2/g)t – Bo = Bo[e(ln2/g)t – 1].
Поскольку в любой популяции имеет место смертность, реальное
значение ее биомассы ко времени t (Bt’) будет меньше рассчитанного по
(11.10). Разница между расчетной (Вt) и реальной (Вt’) ко времени t биомассой продукции популяции соответствует ее элиминации, т.е.
Ве = Вt – Вt’.
60. Продукция коловраток. Коловратки размножаются продуцированием
яиц, число которых в кладке обычно больше одного. По этой причине методы
расчета продукции, разработанные для популяций одноклеточных организмов,
для коловраток не подходят. Для расчета продукции их популяций обычно
применяется так называемый физиологический метод (Методы …, 1968).
Понятие чистой экологической эффективности роста К2 = Р/(P+T),
предложенное первоначально для отдельных особей (гл. 4.2), можно применить
и ко всей популяции. В таком случае Р – продукция популяции, а Т – ее
суммарные траты на дыхание; отсюда К2 является чистой эффективностью
продукции популяции.
Легко доказать, что P = TK2/(1 – K2). У большинства видов значения К2
на организменном уровне снижаются с возрастом – от максимальных – у
новорожденных особей, до нуля к концу жизненного цикла. Однако поскольку
в популяциях коловраток присутствуют особи разных возрастных групп,
популяционные К2, в отличие от К2 для отдельных особей, относительно
постоянны в течение сезона вегетации. У большинства видов значения
популяционных К2 изменяются в пределах 0,2–0,4. Отсюда зная суммарные
траты энергии популяции на дыхание, можно приблизительно определить ее
продукцию.
Пример. Для популяции коловратки Brachionus calyciflorus значения К2
равно 0,33, а ее суммарные траты на дыхание (Т) - 10 ккал·м-3·сутки-1. Тогда
продукция популяции составит 10 · 0,33/(1 – 0,33) = 5 ккал м-3·сутки-1.
61. Поток энергии по трофическим уровням. Рассмотрим превращение
энергии в экосистеме на примере простой пастбищной трофической цепи, в
которой имеется всего три трофических уровня. Первый уровень образуют
травянистые растения, второй – травоядные млекопитающие, например, косули,а третий – хищные млекопитающие, например, волки. Превращения энергии в нашей экосистеме мы выразим в условных «энергетических единицах» (э. е.).
Примем валовую первичную продукцию первого трофического уровня
(ВПП) равной 400 э. е., а затраты растений на дыхание (T) – 50 % от ВПП, т. е. 200 э. е. Тогда чистая первичная продукция (ЧПП) также равна 200 э. е.:
ЧПП = ВПП – D .
100 200 100
Чистую первичную продукцию, образованную растениями, потребляют
травоядные животные, которые составляют второй трофический уровень.
Превращение энергии на нем можно представить в виде балансового равенства,аналогичного энергетическому балансу на организменном уровне (гл. 2.4):ΣRII = ΣPII + ΣTII + ΣNII,
где ΣRII – суммарный рацион всех организмов, составляющий данный уровень;
ΣРII, ΣTII и ΣNII – соответственно их суммарная продукция, траты на дыханиеи неусвоенная часть рациона.
В силу разных причин особи второго трофического уровня способны
потребить лишь некоторую долю ЧПП. Существенная ее часть в виде отмерших органических остатков (подземные части растений, твердая древесина стеблей,ветвей и т.д.) не способна поедаться большинством животных. Она поступает в детритные пищевые цепи и (или) подвергается разложению редуцентами.Однако мы рассмотрим идеальный случай, когда растительноядныемлекопитающие, образующие второй трофический уровень, потребляют всю ЧПП своей экосистемы, т. е. ΣRII = ЧПП = 100 э. е.
Допустим, валовая экологическая эффективность продукции (KIp)
второго трофического уровня равна 0,1, т. е. все гомойотермные организмы
данного уровня расходуют на прирост массы своего тела (ΣРII) 10 %
потребленного ими корма. Остальная часть рациона не или усваивается ими
(ΣHII) или превращается в процессе дыхания в тепловую энергию (ΣТII).
Допустим далее, что ΣТII = ΣHII. Отсюда, превращение энергии на втором
трофическом уровне можно представить в виде:
ΣRII = ΣPII + ΣTII + ΣHII .
100 10 45 45
Величина ΣPII представляет суммарное количество энергии,заключенной в телах травоядных млекопитающих. Только она может быть потреблена хищниками, находящимися на третьем трофическом уровне. В принципе хищники способны потребить всех травоядных животных второго трофического уровня. Поэтому их рацион может составлять вся
величина прироста особей второго трофического уровня, т.е. ΣRIII = ΣРII.
Неусвоенная часть рациона поступает в детритные пищевые цепи.
Допустим, соотношение между различными компонентами баланса
энергии у травоядных и хищных гомойотермных животных одинаково. Тогда
ΣRIII = ΣPIII + ΣTIII + ΣHIII .
10 1 4,5 4,5
Таким образом, в экосистемах с повышением трофического уровня
происходит быстрое уменьшение энергии, накапливаемой в телах живых
организмов. Преобладающая часть энергии, органических соединений,
созданных растениями, в результате дыхания всех живых организмов
экосистемы безвозвратно переходит в тепловую энергию. Однако при дыхании выделяются также углекислый газ и вода, которые вновь возвращаются в экосистему, где могут быть использованы в процессе фотосинтеза.Непереваренные остатки пищи и отмершие организмы всех трофических уровней разлагаются детритофагами и в конечном итоге редуцентами до простых неорганических веществ (вода, углекислый газ, азотистые соединения).Эти вещества вновь могут использоваться продуцентами и редуцентами.Поэтому в экосистемах происходит постоянный круговорот вещества,поскольку одни и те же атомы могут постоянно переходить из одних организмов в другие. Определенная часть продукции экосистемы может выноситься из нее(конечная продукция) или при определенных условиях накапливаться в виде неразалагающихся органических соединений. В предыдущие геологические эпохи (преимущественно в каменноугольный период палеозойской эры) в
биосфере произошло образование органических полезных ископаемых –
каменного угля, нефти, природного газа из слежавшихся мощных слоев остатков наземных растений. В тот период на суше практически не было крупных наземных растительноядных животных. В наше время в экосистемах болот сходным образом происходит образование торфа. Однако чаще всего преобладающая часть чистой первичной продукции, созданной в экосистемах,расходуется в процессах дыхания всех населяющих ее живых организмов и рассеивается в виде тепла.Напротив, в экосистеме имеет место не круговорот, а однонаправленный поток энергии. Электромагнитная энергия солнечного излучения в процессе фотосинтеза преобразуется в энергию химических связей органических веществ.В свою очередь, эта энергия в процессе дыхания всех организмов превращается в тепловую энергию. Последняя, согласно второму закону термодинамики,является самой деградированной формой энергии, которая не способна превращаться в другие формы энергии. Поэтому тепловая энергия,выделяющаяся во внешнюю среду, увеличивает ее энтропию и степень неупорядочненности. Постоянное удаление из экосистемы энтропии,образующейся в результате ее функционирования, по ддерживает е е стабильное состояние в пространстве и во времени.
61Трансформация вещества и энергии в биогеоценозе.
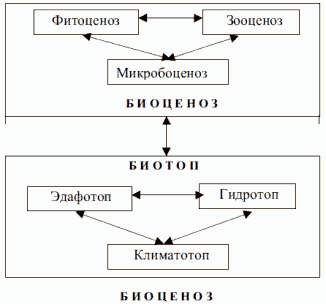 Состав
биогеоценоза можно представить в виде
следующей схемы (рис. 12).
Состав
биогеоценоза можно представить в виде
следующей схемы (рис. 12).
Биогеоценозы являются открытыми системами, которые требуют постоянного поступления вещества и энергии извне. Основным источником этой энергии является солнечный свет, фототрофы улавливают и преобразуют в энергию химических связей синтезируемого органического вещества. Гетеротрофные организмы получают необходимую им энергию вследствие ферментативного разложения органических веществ. Лишь незначительная часть энергии солнечного света, который достигает поверхности Земли (около 1%) фиксируется зелеными растениями, другая же отражается в космос или рассеивается в виде тепла. В свою очередь, организмы, потребляющие зеленые растения, тоже усваивают для построения веществ своего тела лишь незначительную часть энергии химических связей своей пищи (10-20%), а остальные рассеивается в виде тепла, тратясь на процессы жизнедеятельности. То же наблюдается и при поедании фитофагов хищниками и т.п. Поток энергии по трофическим уровням. Рассмотрим превращение энергии в экосистеме на примере простой пастбищной трофической цепи, в которой имеется всего три трофических уровня. Первый уровень образуют травянистые растения, второй - травоядные млекопитающие, например, косули, а третий - хищные млекопитающие, например, волки. Превращения энергии в нашей экосистеме мы выразим в условных «энергетических единицах» (э. е.).
Примем
валовую первичную продукцию первого
трофического уровня (ВПП)
равной 400 э. е., а
затраты растений на дыхание (Т)
- 50 % от ВПП,
т. е. 200 э. е. Тогда
чистая первичная продукция (ЧПП)
также равна 200 э. е.:![]() .
.
Чистую
первичную продукцию, образованную
растениями, потребляют травоядные
животные, которые составляют второй
трофический уровень. Превращение
энергии на нем можно представить в виде
балансового равенства, аналогичного
энергетическому балансу на организменном
уровне (гл. 2.4):![]() где ΣRII
– суммарный рацион всех организмов,
составляющий данный уровень; ΣРII
, ΣTII
и ΣNII
– соответственно их суммарная
продукция, траты на дыхание и неусвоенная
часть рациона. В силу разных причин
особи второго трофического уровня
способны потребить лишь некоторую долю
ЧПП. Существенная ее часть в виде
отмерших органических остатков
(подземные части растений, твердая
древесина стеблей, ветвей и т.д.) не
способна поедаться большинством
животных. Она поступает в детритные
пищевые цепи и (или) подвергается
разложению редуцентами. Однако мы
рассмотрим идеальный случай, когда
растительноядные млекопитающие,
образующие второй трофический уровень,
потребляют всю ЧПП своей экосистемы,
т. е. ΣRII
где ΣRII
– суммарный рацион всех организмов,
составляющий данный уровень; ΣРII
, ΣTII
и ΣNII
– соответственно их суммарная
продукция, траты на дыхание и неусвоенная
часть рациона. В силу разных причин
особи второго трофического уровня
способны потребить лишь некоторую долю
ЧПП. Существенная ее часть в виде
отмерших органических остатков
(подземные части растений, твердая
древесина стеблей, ветвей и т.д.) не
способна поедаться большинством
животных. Она поступает в детритные
пищевые цепи и (или) подвергается
разложению редуцентами. Однако мы
рассмотрим идеальный случай, когда
растительноядные млекопитающие,
образующие второй трофический уровень,
потребляют всю ЧПП своей экосистемы,
т. е. ΣRII
= ЧПП = 100 э. е.
Допустим, валовая экологическая эффективность продукции (KIp) второго трофического уровня равна 0,1, т. е. все гомойотермные организмы данного уровня расходуют на прирост массы своего тела ΣРII 10 % потребленного ими корма. Остальная часть рациона не или усваивается ими ΣHII или превращается в процессе дыхания в тепловую энергию (ΣТII)
Допустим
далее, что ΣТII = ΣHII Отсюда, превращение
энергии на втором трофическом уровне
можно представить в виде:
![]()
62Экологические правила трофических цепей.
Экологические правила трофических уровней. Из разобранных выше закономерностей трансформации энергии в экосистеме следует ряд важных и интересных выводов. А. Число трофических уровней в экосистеме не может быть очень большим. С повышением трофического уровня быстро снижается его продукция, т.е. количество энергии, доступной для использования организмами более высоких трофических уровней. В рассмотренной выше экосистеме из трех трофических уровней (наземные растения – травоядные млекопитающие – хищные млекопитающие) валовая первичная продукция составляет 400 э. е. энергетических единиц, а продукция организмов третьего трофического уровня (хищные млекопитающие) не превышает 1 э. е. Снижение продукции, выражающееся пирамидой продукций, накладывает серьезные ограничения на число трофических уровней. Как правило, в наземных экосистемах, где доминируют гомойтермные организмы (птицы и млекопитающие), число трофических уровней не превышает 3–4. В таких экосистемах второй (первый гетеротрофный) трофический уровень, образован мелкими растительноядными пойкилотермными животными, преимущественно насекомыми и их личинками. При этом трофическая цепь принимает следующий вид : [I] растения → [II] членистоногие → [III] насекомоядные птицы и млекопитающие → [IV] хищные птицы и млекопитающие. В экосистемах, где гетеротрофные уровни пищевых цепей (за исключением последнего) формируют пойкилотермные организмы, число трофических уровней может возрастать до 6–7. Подобная ситуация часто имеет место в водных экосистемах. Приведем пример подобной цепи : [I] Планктонные автотрофные прокариоты и протисты → [II] планктонные гетеротрофные протисты - амебы, инфузории → [III] мелкие планктонные беспозвоночные с фильтрационным типом питания (ветвистоусые рачки, коловратки) → [IV] хищные планктонные беспозвоночные (веслоногие рачки) → [V] планктоноядные рыбы (плотва, уклея и т. п.) → [VI] хищные рыбы (щука, окунь) → [VII] крупные рыбоядные хищники (выдра, рыбоядные птицы, а также человек). Последний трофический уровень в данном случае образуют уже наземные гомойотермные организмы, т.е. пищевые цепи выходят из водных экосистем в наземные. Более длинные пищевые цепи в сообществах пойкилотермных организмов обусловлены тем, значения коэффициентов валовой экологической эффективности продукции (К ) у них значительно выше, чем у 1гомойотермных организмов. Если максимальные значения К у гомойотермных 1животных не превышает 0,1 (обычно– не более 0,01–0,05, то у пойкилотермных К1 часто достигает 0,3–0,40 и даже более. Отсюда пойкилотермные организмы способны передать на последующий трофический уровень приблизительно в три и более раз больше энергии потребляемой ими пищи, чем гомойотермные. При этом пойкилотермные животные вследствие своих мелких размеров способны гораздо более полно использовать пищевые ресурсы предыдущего трофического уровня. ическая эффективность превращения энергии в трофических цепях.
63 Экологические пирамиды. В результате трофических взаимодействий особей, образующих экосистему, в ней формируется определенная трофическая структура. Ее можно выразить в виде экологических пирамид, основанием которых является первый трофический уровень (уровень продуцентов), а последующие этажи и вершину образуют последующие уровни. Экологические пирамиды можно отнести к трем основным типам:
пирамиды численности, которые отражают общую численность организмов каждого трофического уровня;
пирамиды биомасс, характеризующих общую массу особей каждого трофического уровня;
пирамиды продукции, характеризующие продукцию каждого трофического уровня.
Пирамиды численности, как правило, наименее информативны и показательны, поскольку численность организмов одного трофического уровня в экосистеме в значительной степени зависит от их размеров. Например, масса одной лисицы равна массе нескольких сотен мышей.
Обычно численность гетеротрофных организмов в экосистеме выше, чем автотрофных. На одном дереве (первый трофический уровень) может кормиться до нескольких тысяч насекомых (второй трофический уровень). С повышением трофического уровня гетеротрофных организмов средние размеры особей, находящихся на нем, обычно возрастают, а их численность снижается. Поэтому пирамиды численности в экосистемах часто имеют вид «новогодней елки».
Пирамиды биомасс гораздо лучше выражают соотношения между разными трофическими уровнями экосистемы. В целом, биомасса более низких уровней превышает биомассу более высоких. Однако из этого правила имеются существенные исключения. Например, в морях биомасса сообществ растительноядного зоопланктона существенно (иногда в 2-3 раза) больше биомассы фитопланктона, представленного преимущественно одноклеточными водорослями. Это объясняется тем, что водоросли очень быстро выедаются зоопланктоном, но от полного выедания их предохраняет очень высокая скорость деления клеток.
В целом пирамиды численности приводят к переоценке значения мелких организмов, а пирамиды биомасс - к переоценке роли крупных организмов. Поэтому наиболее полное представление о функциональной организации экосистем дают пирамиды продукций. При этом продукцию каждого трофического уровня лучше представлять в единых единицах измерения, лучше всего в энергетических единицах. В таком случае пирамиды продукций станут пирамидами энергий.
В противоположность пирамидам численности и биомассы, отражающим статику системы (т. е. характеризующим численность и массу организмов в данный момент времени), пирамиды продукции характеризуют скорости прохождения энергии пищи по трофическим цепям. Если правильно учтены все величины поступления и расхода энергии в трофической цепи, то в соответствии со вторым законом термодинамики пирамиды продукции всегда будут иметь правильную форму.
Численность и биомасса организмов, которые может поддерживать какой- либо уровень в тех или иных экологических условиях, зависит не от количества фиксированной энергии, имеющейся в данный момент на предыдущем уровне (т. е. от биомассы последнего), а от скорости продуцирования пищи на нем.
Одним из важнейших показателей продукционных возможностей организмов является среднесуточный Р/В-коэффициент, или удельная продукция. С увеличением массы организмов их Р/В-коэффициенты быстро снижаются (Заика, 1983). С повышением трофического уровня происходит быстрое снижение продукции организмов и их Р/В-коэффициентов. Это в основном обусловлено тем, что средние размеры гетеротрофных организмов на последующем трофических уровнях выше, чем на предыдущем.