

17. Активный обмен у животных и методы его определения.
Интенсивность потребления кислорода у всех живых организмов возрастает при любых формах их физической активности, включая передвижение (плавание, бег, машущий полет), добывание пищи, заботу о потомстве и пр. В этой связи выделяют ряд уровней энергетического обмена особей в зависимости от степени их
двигательной активности.
Основной обмен (basal metabolism) характеризует минимально
возможный уровень обмена у особей с данной массой тела. У гомойотермных
животных он измеряется в условиях полного покоя, например, во время сна
при температуре критической точки или в термонейтральной зоне. Энергия,
выделяющаяся при дыхании, в этом случае расходуется только на
функционирование внутренних органов – дыхательной, пищеварительной,
нервной и других систем организма.
Стандартный обмен имеет место при минимальной двигательной
активности организма. Поэтому его часто называют обменом покоя (resting
metabolism). Большинство данных по скорости обмена у животных получены
при их содержании в различных экспериментальных установках, т. е. в
условиях весьма ограниченной двигательной активности. Поэтому полученные
результаты, использованные затем для расчетов уравнений (3.1),
представленных в табл. 3.2, характеризуют уровень их обмена, близкий к
стандартному.
Активным обменом (active metabolism) называют уровень обмена особи
при более-менее существенном уровне ее двигательной активности. Он
является суммой стандартного обмена и затрат энергии на передвижении. У
ряда видов млекопитающих с разной массой тела интенсивность активного
обмена (Qact/W) при средних скоростях движения возрастает прямо
пропорционально скорости их бега (V).
Отсюда зависимость Qact/W от V у млекопитающих в упрощенном виде может быть выражена уравнением:
Qact/W = f + gV. (3.9)
18. Зависимость интенсивность дыхания у пойкилотермных и гомойотермных животных от температуры.
Наскорость дыхания организмов, как и на другие их жизненные процессы,
значительное воздействие оказывает температура среды. Поскольку масса
особей также влияет на скорость дыхания, воздействие температуры на этот
показатель следует изучать на одноразмерных особях.
Характер температурной зависимости скорости (Q) или интенсивности
дыхания (Q/W) у пойкилотермных и гомойтермных животных резко
различается
(рис. 3.2).
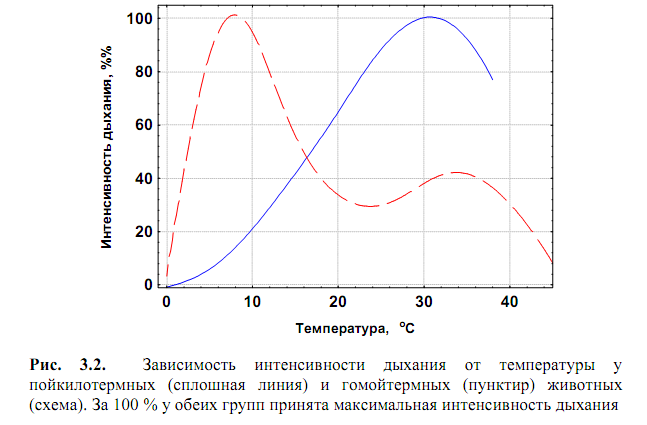
У пойкилотермных животных, температура тела которых равна или очень
мало отличается от температуры среды, значения Q или Q/W с температурой
возрастают, а Q10 для них снижаются. Примером этого являются приводимые
уже данные для черноморского моллюска Modiola adriatica (рис. 1.2). При
изучении изменений Q/W у колорадского жука в широком температурном
диапазоне выявлена аналогичная закономерность (табл. 3.2).
Заметим, что значения ΔF для Q/W у этого вида остаются практически
постоянными в интервале 7–20 оС, но быстро снижаются при дальнейшем
повышении температуры. Следовательно, температурная зависимость Q/W у колорадского жука может быть описана уравнением Вант-Гоффа – Аррениуса
только в температурном диапазоне 7–20 оС.
Однако возрастание Q (или Q/W) с температурой у пойкилотермных
животных имеет место лишь в пределах оптимальных температур, понимаемых
здесь в широком смысле, включая зоны оптимума и нормы на кривой
толерантности Шелфорда. При дальнейшем повышении температуры за
пределы этой зоны скорость метаболизма снижается. Проиллюстрируем это на
примере эвритермного легочного моллюска Lymnaea hoduthae (Хмелева,
Голубев, Лаенко, 1985). Данный вид обитает в геотермальном источнике
Ходутка (Камчатка) в интервале температур от 5–7 до 35 оС, но не теряет
жизнеспособности в течение нескольких часов при ее повышении до 40 оС,
однако размножается в более узком интервале – от 12 до 33 оС.
Тем не менее, характер изменений Q/W с температурой в обоих случаях
одинаков. Этот показатель вначале возрастает с температурой, достигает
максимума при определенных ее значениях вблизи верхнего предела зоны
температурного оптимума, а затем резко снижается. Значения Q10 для Q/W в
целом снижаются с температурой, становясь в сублетальной зоне меньше
единицы (рис. 3.3б).
Снижение Q/W при повышенных температурах часто сопровождается
ростом смертности особей. Поэтому характер температурной зависимости
Q/W может являться одним из критериев нахождения верхней температурной
границы зоны нормы.
У гомойотермных животных (рис. 3.2) на характер температурной
зависимости скорости метаболизма значительное воздействие оказывает
необходимость поддержания постоянной температуры тела при изменениях
температуры среды. Если температура их тела выше температуры среды (а
чаще всего так и бывает), часть образующегося в процессе метаболизма тепларассеивается в среде. Согласно закону остывания Ньютона, величина
теплоотдачи (Н) сплошного нагретого тела (шара) равна произведению
удельной теплопроводности тела (C), площади его поверхности (S) и разности температуры поверхности тела (Тт) и среды (Тс), т. е.
Н = CS(Tт – Тс). (3.7)
Ряд ученых, начиная с М. Рубнера (1882), считает возможным
использовать уравнение (3.7) для характеристики величин потерь
метаболического тепла у гомойотермных животных. В частности, оно вполне подходит для человека, у которого очень слабо развиты волосяной покров и
подкожная жировая прослойка, выполняющие теплоизолирующую функцию.
Однако для большинства птиц и млекопитающих, имеющих хорошо
развитые внешние покровы (мех, перья и т. п.) или толстый слой подкожного
жира, более подходит закон Фурье, описывающий теплоотдачу тел, покрытых теплоизолирующей оболочкой:
Н = hS(Tт –Тс)/L, (3.8)
где h – удельная теплопроводность внешних покровов, L – их толщина, S –
площадь поверхности тела (Дольник, 2002).
Из (3.7) и (3.8) следует, что чем выше разность Tт–Тс, тем большее
количество тепла (при прочих равных условиях) передается от нагретого тела
во внешнюю среду. Величина Тс у отдельных видов достаточно постоянна; для всей поверхности тела человека, например, она составляет в среднем около 25-27оС. Поэтому чем ниже температура среды, чем больше тепла теряет организм, тем выше должен быть уровень его метаболизма для компенсации потерь тепла.
Поэтому максимальный уровень метаболизма у гомойотермных
животных («вершинный обмен») отмечается при некоторой температуре,
близкой к нижнему пределу зоны ее толерантных значений («температура
холодового оцепенения»). При дальнейшем повышении температуры скорость метаболизма снижается, достигая некоторого минимального значения («основной обмен») при определенной температуре среды («критическая точка») или в узком температурном интервале («термонейтральная зона»). «Критическая точка» равна или очень близка к температуре внешних покровов животного, поэтому при ней между организмом и средой устанавливается тепловое равновесие и теплоотдача организма во внешнюю среду становится равной нулю. Например, у человека минимальный уровень обмена наблюдается в узком температурном интервале 25–27 оС, практически совпадающим со средней температурой поверхности его тела.
При температуре среды выше критической точки тепло из внешней среды
поступает в организм, что ведет к его перегреву. Это делает необходимым
отведение излишнего тепла из организма, что достигается посредством
усиленного потоотделения, учащения дыхания, увеличения поверхностного
кровотока. На эти процессы расходуется дополнительная энергия, что приводит к некоторому повышению уровню метаболизма. При дальнейшем повышении температуры уровень метаболизма быстро снижается, становясь равным нулю при температуре смерти от перегрева (Слоним, 1971).