

5. Основные свойства белковых фракций крови и значение их определения для диагностики. Методы исследования. Эмбриоспецифические белки и их значение. Энзимодиагностика.
Белки плазмы крови: образуют буферную систему крови и поддерживают рН крови в пределах 7,37-7,43, поддерживают осмотическое давление, удерживая воду в сосудистом русле, транспортируют метаболиты , витамины, ионы металлов , лекарства, определяют вязкость крови, являются резервом аминокислот для организма, выполняют защитную роль. Эмбриоспецифические белки - их синтез репрессирован в организме взрослого человека. Результаты поиска эмбриоспецифических белков в сыворотке крови больных с различной патологией позволили предложить иммунохимический тест на первичный рак печени. Энзимодиагностика - методы диагностики болезней, патологических состояний и процессов, основанные на определении активности энзимов (ферментов) в биологических жидкостях.
6. Глобулярные и фибриллярные белки, простые и сложные. Представления о структуре фибриллярных белков. Коллаген как основной белок соединительной ткани.
Если белки состоят только из аминокислот, их называют простыми. Если белки содержат помимо аминокислот еще и неаминокислотный компонент (простетическую группу), их называют сложными. Если пептидные цепи уложены в виде клубка, то такие белки называются глобулярными. Если полипептидные цепи уложены в пучки нитей, они носят название фибриллярных белков. Фибриллярные белки — белки, имеющие вытянутую нитевидную структуру, в которой отношение длинной оси молекулы к короткой составляет от 80 до 150. Большинство фибриллярных белков не растворяется в воде, имеет большую молекулярную массу и высоко регулярную пространственную структуру, которая стабилизируется, главным образом, взаимодействиями между различными полипептидными цепями. Коллаген — фибриллярный белок, составляющий основу соединительной ткани организма (сухожилие, кость, хрящ, дерма) и обеспечивающий её прочность и эластичность. Молекула коллагена представляет собой правозакрученную спираль из трёх α-цепей. Продуктом денатурации коллагена является желатин
7. Хромопротеины, важнейшие представители, строение и роль в организме. Типы гемоглобинов и их изменение в процессе онтогенеза. Гемоглобинопатии.
Хромопротеины - сложные белки, состоящие из простого белка и связанного с ним окрашенного небелкового компонента — простетической группы. Различают гемопротеины, магнийпорфирины и флавопротеины. Хромопротеины участвуют в процессах жизнедеятельности: как фотосинтез, клеточное дыхание, дыхание всего организма, транспорт кислорода и углекислого газа, окислительно-восстановительные реакции, свето- и цветовосприятие. Хромопротеины являются важнейшими участниками аккумулирования энергии. Нормальные гемоглобины - примитивный гемоглобин (гем. Р), фетальный гемоглобин (гем. F), гемоглобин взрослых (гем. А); аномальные - гемоглобин S,C,D,E. В процессе онтогенеза изменяется не только количественное содержание гемоглобина, но и его качественная характеристика. Гемоглобин плода и взрослого организма отличается качественно по кривой диссоциации, устойчивостью по отношению к денатурации, скоростью реакции трансгемирования и т. д. В процессе онтогенеза процентное содержание «фетального» гемоглобина падает. Но и в крови взрослого может содержаться небольшой процент «фетального» гемоглобина. Гемоглобинопатия — наследственное или врождённое изменение или нарушение структуры белка гемоглобина, обычно приводящее к клинически или лабораторно наблюдаемым изменениям в его кислород-транспортирующей функции либо в строении и функции эритроцитов.
8. Строение нуклеиновых кислот. Азотистые основания и сахара, входящие в состав ДНК и РНК. Нуклеозиды и нуклеотиды. Адениловые динуклеотиды (НАД, НАДФ, ФАД). Связи, формирующие первичную структуру ДНК и РНК, –5`-фосфатный и 3`- гидроксильный концы полинуклеотидных цепей. Вторичная структура ДНК и РНК. Денатурация и ренатурация ДНК.
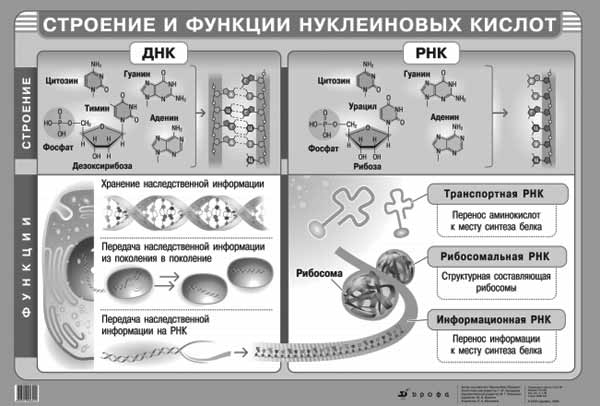
К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C), которые входят в состав как ДНК, так и РНК. Тимин (T) входит в состав только ДНК, а урацил (U) встречается только в РНК. Сахара: ДНК - дезоксирибоза, РНК - рибоза. Нуклеотиды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Нуклеозиды — это гликозиламины, содержащие азотистое основание, связанное с сахаром (рибозой или дезоксирибозой).
Никотинамидадениндинуклеотиид — кофермент, присутствующий во всех живых клетках, входит в состав ферментов группы дегидрогеназ.
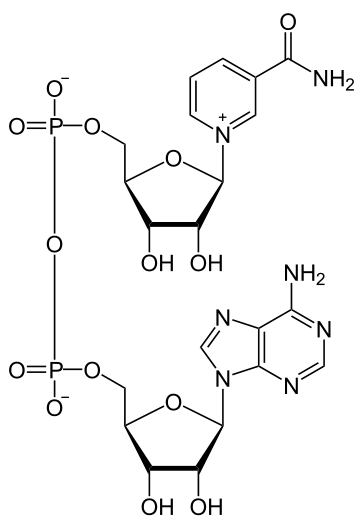
Флавинадениндинуклеотид — кофермент, принимающий участие во многих окислительно-восстановительных биохимических процессах.

Никотинамидадениндинуклеотидфосфат — кофермент некоторых дегидрогеназ.
Вторичной структуры ДНК является двойная спираль. Эта структура образуется из двух взаимно комплементарных антипараллельных полидезоксирибонуклеотидных цепей, закрученных относительно друг друга и общей оси в правую спираль. При этом азотистые основания обращены внутрь двойной спирали, а сахарофосфатный остов — наружу. Молекулы РНК представляют собой единичные полинуклеотидные цепи. Отдельные участки молекулы РНК могут соединяться и образовывать двойные спирали. Спаривание оснований в таких спиралях бывает неполным. В результате внутримолекулярного спаривания оснований формируются такие вторичные структуры, как стебель-петля. Водородные связи и межплоскостные взаимодействия, стабилизирующие двойную спираль слабые, и при небольших воздействиях происходит разделение цепей — денатурация. Двухцепочечная спиральная ДНК в растворе легко разрушается при нагревании до температур, близких к 100°С. Процесс воссоединения, называемый ренатурацией, происходит при понижении температуры или рН.