

40. Зависимость скорости ферментативной реакции от концентрации субстрата. Графики Михаэлиса-Ментен и Лайнуивера-Бэрка. Медико-биологическое значение константы Михаэлиса.
Число молекул субстрата, претерпивающих изменение в течение минуты, в расчете на единицу веса фермента при оптимальных условиях его действия называется молекулярной активностью фермента. При увеличении количества ферментов скорость ферментативной реакции повышается до некоторого предела, который характеризуется количеством субстрата, доступным действию фермента. В то же время, при постоянной концентрации фермента увеличение количества субстрата приводит вначале к быстрому, затем к более медленному росту скорости ферментативной реакции, пока не достигается максимальная скорость, остающаяся практически неизменной при дальнейшем увеличении концентрации субстрата. ЗСФРОКС: V=Vmax/1+Km/S, где V - скорость ферментативной реакции, Vmax - максимальная скорость ее при бесконечно большой концентрации субстрата, S - концентрация субстрата в моль/л, Кm - константа Михаэлиса.
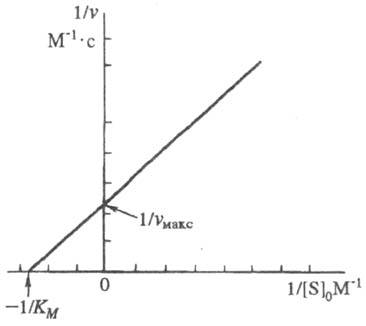 Величина
Км численно равна концентрации субстрата,
при которой скорость р-ции равна 1/2 Vмакс
, поэтому Км часто служит мерой сродства
субстрата и фермента, однако это
справедливо лишь, если Км ~ Ks.
Величина
Км численно равна концентрации субстрата,
при которой скорость р-ции равна 1/2 Vмакс
, поэтому Км часто служит мерой сродства
субстрата и фермента, однако это
справедливо лишь, если Км ~ Ks.
Она соответствует концентрации субстрата, при которой скорость реакции равна половине максимальной и служит мерой химического сродства меду ферментом и субстратом, мерой их способности образовывать фермент-субстратный комплекс.
41. Зависимость скорости ферментативной реакции от концентрации субстрата. График Михаэлиса-Ментен. Медико-биологическое значение константы Михаэлиса.
Число молекул субстрата, претерпивающих изменение в течение минуты, в расчете на единицу веса фермента при оптимальных условиях его действия называется молекулярной активностью фермента. При увеличении количества ферментов скорость ферментативной реакции повышается до некоторого предела, который характеризуется количеством субстрата, доступным действию фермента. В то же время, при постоянной концентрации фермента увеличение количества субстрата приводит вначале к быстрому, затем к более медленному росту скорости ферментативной реакции, пока не достигается максимальная скорость, остающаяся практически неизменной при дальнейшем увеличении концентрации субстрата. ЗСФРОКС: V=Vmax/1+Km/S, где V - скорость ферментативной реакции, Vmax - максимальная скорость ее при бесконечно большой концентрации субстрата, S - концентрация субстрата в моль/л, Кm - константа Михаэлиса.
Величина Км численно равна концентрации субстрата, при которой скорость р-ции равна 1/2 Vмакс , поэтому Км часто служит мерой сродства субстрата и фермента, однако это справедливо лишь, если Км ~ Ks.
Она соответствует концентрации субстрата, при которой скорость реакции равна половине максимальной и служит мерой химического сродства меду ферментом и субстратом, мерой их способности образовывать фермент-субстратный комплекс.
42. Регуляция активности ферментов. Различные способы активации и ингибирования ферментов.
Факторы, регулирующие активность ферментов, разнообразны по своей природе. Физические факторы (температура, давление, свет, магнитное поле, электрические импульсы) оказывают менее специфическое действие, чем химические. В свою очередь действие последних также может быть разделено на несколько типов. Одни химические вещества связываются с активным центром фермента, например субстраты, кофакторы, конкурентные ингибиторы, что приводит к изменению ферментативной активности. Другие вещества взаимодействуют со специальными участками на поверхности молекулы определенного типа фермента, не имеющими непосредственного отношения к центрам каталитической активности, но тем не менее приводящими к ее изменению. Активаторы органической природы: желчные кислоты (активируют поджелудочную липазу), энтерокиназа (активирует трипсиноген), глутатион, цистеин, витамин С (повышают активность оскидоредуктаз).
Активаторы неорганической природы: например, HCl активирует пепсиноген, ионы ме таллов (Na, Cl, K, Mg, Mn, Zn). Если ингибитор связывается с молекулой фермента слабыми связями, то такой ингибитор легко удаляется и активность фермента восстанавливается. Если ингибитор связывается с молекулой фермента прочными ковалентными связями, то наступает необратимое подавление активности фермента.
43. Регуляция действия ферментов, аллостерические механизмы, фосфорилирование - дефосфорилирование. Примеры метаболических путей, регулируемых этими механизмами. Классификация ферментов, ее принципы. Характеристика оксидоредуктаз.
В живой клетке скорость ферментативных реакций находится под строгим контролем, что позволяет каждой метаболической цепочке реакций постоянно изменяться, приспосабливаясь к меняющимся потребностям клетки в продукте. В каждой метаболической цепи есть фермент, который задает скорость всей цепочке реакций. Он называется регуляторным ферментом. Существует несколько способов регуляции действия ферментов: изменение активности фермента при его постоянной концентрации; изменение концентрации фермента, обычно в результате ускорения (индукции) или торможения (репрессии) синтеза фермента. Аллостерическим механизмом регуляции активности ферментов называют регуляторный механизм, в котором контроль активности фермента реализуется путем изменения конформации белковой молекулы, индуцируемого связыванием метаболита-регулятора в особом (аллостерическом) центре, пространственно удаленном от активного центра. Фосфорилирование — процесс переноса остатка фосфорной кислоты от фосфорилирующего агента- донора к субстрату, как правило, катализируемый ферментами и ведущий к образованию эфиров фосфорной кислоты. Дефосфорилирование - отщепление остатка фосфорной кислоты от молекулы фосфорсодержащего соединения. КФ 1: Оксидоредуктазы, КФ 2: Трансферазы, КФ 3: Гидролазы, КФ 4: Лиазы, КФ 5: Изомеразы, КФ 6: Лигазы. Оксидоредуктазы, катализирующие окисление или восстановление. Пример: каталаза, алкогольдегидрогеназа.