

Класс ксантотриховые
XANTHOTRICHOPHYCEAE
Представитель – Tribonema sp. (трибонема). Обитает в пресных водоемах. Имеет нитчатый (трихальный) таллом (рис. 62).
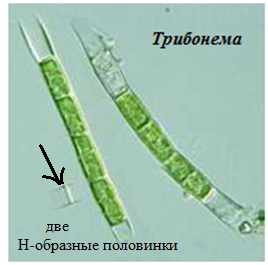
Рис. 62. Трибонема.
В начале жизни трибонема прикрепляется к субстрату с помощью базальной клетки, которая впоследствии отмирает, и водоросль всплывает на поверхность водоема, часто образуя тину. Клетки одноядерные, с желтовато-зелеными хроматофорами, расположенными постенно. Бесполое размножение осуществляется при помощи разножгутиковых зооспор, формирующихся в вегетативных клетках таллома. В окружающую среду зооспоры выходят через створки клеток. Разрыв нитей происходит в середине клеток, после чего трибонему легко отличить от других нитчатых водорослей по характерному очертанию концов ее нитей в виде двух Н-образных половин. При наступлении неблагоприятных условий в нитях трибонемы образуются апланоспоры с толстой двустворчатой оболочкой, заполненные каплями масла и жира.
Класс ксантосифоновые
XANTHOSIPHONOPHYCEAE
Представитель – Botrydium sp. (ботридий). Обитает на влажной почве около воды в виде желто-зеленых шариков, диаметром до 1 мм, напоминающих рассыпанный бисер (рис. 63). Каждый такой шарик – клетка ботридиума. Книзу клетка постепенно сужается и переходит в ветвящиеся бесцветные ризоиды, погруженные в почву. В центре клетки имеется вакуоль, в пристенном слое цитоплазмы находятся мелкие ядра, многочисленные хроматофоры и капли масла. Такой тип организации таллома является сифональным, или неклеточным. Зооспоры массово образуются во время дождя и выходят в окружающую среду через отверстие на вершине таллома, напоминая извержение вулкана.
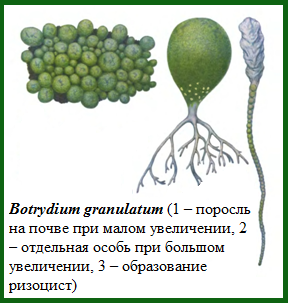
Рис. 63.Ботридий.
Представитель – Vaucheria sp. (вошерия). Названа в честь швейцарского ботаника Жана Пьера Этьена Воше, который сам описал её (рис. 64).

Рис. 64. Вошерия.
В природных условиях около 60 видов этого рода представляют собой крупные слаборазветвленные нити, образующие грязно-зеленые ватообразные скопления в воде или обширные дерновины темно-зеленого цвета на влажной земле, прикрепляясь к почве с помощью ризоидов. Таллом сифональный (без клеточных перегородок, т.е. как бы одна вытянутая многоядерная клетка, с многочисленными мелкими хроматофорами дисковидной формы), ветвистый. Хроматофоры расположены постенно, пиреноиды отсутствуют.
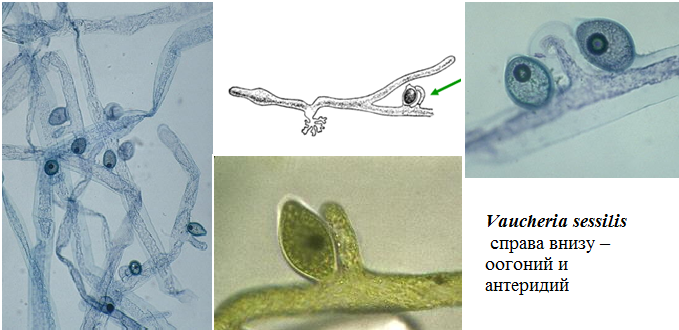
Рис. 65. Оогоний и антеридий Вошерии.
При бесполом размножении на конце нити образуется зооспорангий с одной большой зооспорой, покрытой по периферии большим числом попарно расположенных гладких жгутиков (это происходит за счет того, что образующиеся в зооспорангии зооспоры не расходятся, соединяясь вместе, поэтому этот комплекс носит название синзооспора). Зооспора некоторое время плавает в воде, а затем сбрасывает жгутики и прорастает в новый таллом. Половой процесс – оогамия. Антеридии и оогонии одноклеточные (рис. 65). После периода покоя зигота делится путем мейоза и прорастает, образуя новую особь.
☼ Отдел ДИНОФИТОВЫЕ, ДИНОФЛАГЕЛЛЯТЫ DINOPHYTA
Динофитовые и криптофитовые водоросли, которых до недавнего времени объединяли в единый отдел пиррофитовых водорослей – древние организмы. Вероятно, они произошли от примитивных амебоидных видов, что роднит их с золотистыми водорослями, а особенности пигментного состава указывают на их родство с бурыми водорослями. Достоверны находки панцирей динофитовых водорослей (рис. 66) в отложениях, возраст которых – около 250-280 млн. лет.
Численность: описано 5—6 тысяч видов динофлагеллят.
Среда обитания: преимущественно в чистых пресных и соленых водах (рис. 67). Известны криофильные формы, обитающие в снегу.
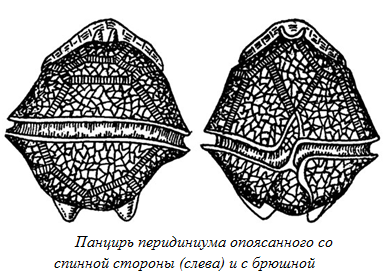
Рис. 66. Панцирь перидиниума опоясанного со спинной стороны (слева) и брюшной - справа.
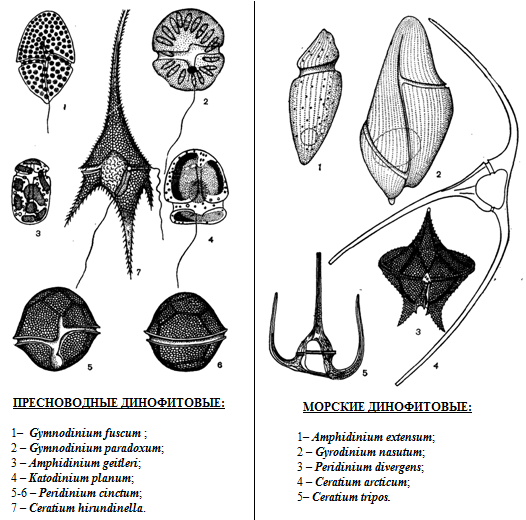
Рис. 67. Динофитовые: слева – пресноводные: 1 – Gymnodinium fuscum, 2 – Gymnodinium paradoxum, 3 – Amrhydinium geitleri, 4 – Katodinium planum, 5-6 – Peridinium cinctum, 7 – Ceratium hirundinella, справа – морские: 1 – Amphidinium extensum, 2 – Dyrodinium nasutum, 3 – Peridinium divergens, 4 – Ceratium arcticum, 5 – Ceratium tripos.
Уровень организации: отдел объединяет представителей, имеющих, в основном, одноклеточные организмы с двумя гетероконтными и гетероморфными жгутиками. Расположенный вдоль продольной оси клетки жгутик сообщает ей поступательное движение, второй, перпендикулярный первому, – вращательное движение.
Структура талломов представлена 5 типами (рис. 68):
1) преимущественно монадная, но есть
2) амебоидные, пальмеллоидные, а также
3) коккоидные и нитчатые у неподвижных видов.

Рис. 68. Основные типы структуры талломов у Динофитовых: 1 – амебоидная (Dinamoedium varians); 2 – монадная (Hemidinium nasutum); 3-4 – пальмеллоидная (Cystodinium montanum); 5-7 – коккоидная (Cystodinium steinii, Stylodinium sphaera, Tetradinium intermedium); 8 – нитчатая (Dinothrix paradoxa).
Строение клетки. Клетки динофитовых водорослей имеют разное строение. Самые примитивные из них – амебоидные – лишены плотной оболочки и, следовательно, не имеют постоянной формы. На клетке легко возникают и также легко исчезают различные выросты (ризоподии), благодаря которым эти водоросли способны передвигаться ползком, как простейшие животные – амебы.
Особенно широко представлены в этом отделе монады, имеющие плотную клеточную стенку (теку) – целлюлозный панцирь, который состоит из щитков, соединенных швами. Два жгутика располагаются в особых образованиях – бороздах; в поперечной борозде лежит уплощенный лентовидный жгутик, придающий клетке вращательное движение, а в продольной борозде – нитевидный жгутик, который определяет поступательное движение клетки. Особенностью динофитовых водорослей является строение ядра. Оно одно, крупное; похоже на наследственный аппарат сине-зеленых водорослей, но имеет оболочку. В хроматофорах содержатся пигменты хлорофилл а и с, каротиноиды, ксантофиллы, близкий к фукоксантину – перидин. Запасной продукт – крахмал и жир.
Питание у большинства динофлагеллят миксотрофное (наряду с автотрофным наблюдается и гетеротрофное).
Размножаются 1) делением клетки;
2) реже с помощью зооспор (с таким же расположением жгутиков, как и у вегетативной клетки);
3) половой процесс (изогамия) известен лишь у единичных представителей.
Цистообразование. У динофитовых водорослей могут появляться особые толстостенные образования (цисты), благодаря которым особь сохраняет жизнеспособность в течение многих лет.
Роль в природе, жизни человека и в биогеоценозах. Массовое развитие динофитовых водорослей вызывает «цветение» воды, приводящее к гибели обитателей водоемов вследствие выделения сакситоксина. Многие динофитовые входят в симбиоз с сине-зелеными водорослями, медузами, кораллами и т.д.
Динофитовые водоросли, как и криптофитовые, очень чувствительны к изменениям внешней среды, особенно к органическому загрязнению. Это позволяет применять некоторые виды отдела в качестве тест-объектов при санитарно-биологическом анализе воды. Многие виды активно участвуют в процессе самоочищения загрязненных промышленными и бытовыми стоками вод. Развитие их в большом количестве может служить надежным показателем чистоты воды.
Кроме того, динофитовые водоросли используют в геологии для определения возраста осадочных пород (их панцири хорошо сохраняются в ископаемом состоянии).
☼ Отдел КРИПТОФИТОВЫЕ, или КРИПТОМОНАДЫ CRYPTOPHYTA
Криптофиты – небольшая, но экологически и эволюционно очень важная группа. Положение криптомонад среди водорослей до сих пор дискутируется. А. Пашер полагал, что они произошли от хризомонад, и от криптомонад выводил перидинеи. Он объединил криптомонады и перидинеи в отдел Pirrophyta. Большинство альгологов не соглашается с таким объдинением и исключает криптофитовых из пиррофитовых, рассматривая их как самостоятельную группу на основании того, что криптофиты отличаются от динофит строением ядра, жгутикового аппарата, составом пигментов, биохимическими особенностями. Все эти данные свидетельствуют против их близкого филогенетического родства.
Численность: к настоящему времени описано более 200 видов криптофитовых водорослей (приблизительно 100 пресноводных и 100 морских), включенных в класс Cryptophyceae и два порядка – Cryptomonadales и Goniomonadales.
Среда обитания: криптофиты – водоросли морских и континентальных вод, преимущественно обитающие в широком температурном диапазоне. Пресноводные представители предпочитают искусственные и естественные водоёмы со стоячей водой. Среди криптофит встречаются типичные сфагнофиллы, живущие в болотах с низким значением рН. Они отсутствуют в горячих источниках и гиперсолёных водоёмах.
Уровень организации: почти все криптофиты – одноклеточные подвижные жгутиконосцы. На переднем конце клеток расположены два жгутика, длина которых сопоставима с длиной клетки. При движении они или оба направлены вперёд, или один – вперёд, а другой – назад.
Структура таллома: монадная, только 1 род образует нитчатый таллом и некоторые стадии пальмеллоидные.
Строение клетки: обычно клетки уплощённые, их форма разнообразная. От переднего края отходит продольная борозда, которая не доходит до заднего конца тела (рис. 69).
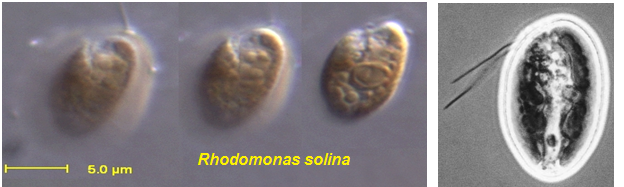
Рис. 69. Родомонас.
Есть мешковидная глотка, по её краю располагаются трихоцисты – стрекательные структуры, окруженные мембраной и содержащие 2 ленты, скрученные рулоном наподобие цилиндров. Более крупный цилиндр состоит из намотанного по спирали лентовидного мембранного материала, в его углублении вверху находится второй цилиндр. При химическом и физическом раздражении туго скрученные ленты распрямляются, разрывая мембрану трихоцисты и плазмалемму клетки, сначала выбрасывается маленький цилиндр, вытягивая за собой большой. По периферии клетки также могут располагаться трихоцисты, но более мелких размеров. На переднем конце тела расположены сократительные вакуоли, изливающие своё содержимое в глотку или борозду. Единственное ядро находится в задней части клетки.
У криптофит клеточные покровы представлены плазмалеммой, над которой могут располагаться пластинки, чешуйки, слизь или их комбинация.
Стигма встречается не у всех криптомонад. Светочувствительный глазок связан с хлоропластом, но не связан со жгутиковым аппаратом, поэтому может располагаться и в центре клетки. Он состоит из большого числа пигментных глобул.
В клетках криптомонад расположены один или два хлоропласта, окрашенные в разные цвета – от сине-зелёного и оливково-зелёного до жёлто-бурого, коричневого и тёмно-красного. Оболочка хлоропласта состоит из четырёх мембран. В хлоропластах могут быть пиреноиды. Иногда хлоропласты отсутствуют.
В хлоропластах присутствует ряд пигментов: помимо хлорофилла а и с, криптофикоциан и криптофикоэритрин. Каждая фотосинтезирующая криптомонада имеет только один тип фикобилипротеина – или фикоэритрин, или фикоцианин, но никогда вместе. Криптомонады образуют несколько уникальных ксантофиллов, таких, как аллоксантин. У них встречаются α- и β-каротины, крококсантин, зеаксантин и монадоксантин.
Основное запасаемое вещество у криптофит – крахмал. В цитоплазме могут присутствовать капли липидов.
Питание: среди криптомонад встречаются автотрофы, гетеротрофы (сапротрофы и фаготрофы) и миксотрофы.
Размножение: 1) основной способ размножения криптомонад – вегетативный, за счёт деления клетки пополам с помощью борозды деления.
2) лишь у некоторых известен половой процесс (хологамия).
Цистообразование: покоящиеся цисты окружены толстой оболочкой.
Роль в природе, жизни человека и в биогеоценозах. В водоёмах криптофиты играют значительную роль в круговороте кислорода, углерода, азота и фосфора, в синтезе органического вещества из минеральных веществ. Криптофитовые – типичные представители планктона, изредка они встречаются в иле солёных озёр и среди детрита в пресных водоёмах.
Криптофиты играют важную роль в холодных водах, особенно зимой и ранней весной. Например, они могут доминировать в весеннем цветении фитопланктона в Северном море, где, возможно, являются важным звеном в пищевых цепях. В покрытых льдами озёрах Антарктиды виды криптомонад могут доминировать в альгофлоре, давая более 70% биомассы фитопланктона.
Бесцветные представители распространены в водах, загрязнённых органическими веществами, их много в сточной воде, поэтому они могут служить показателем загрязнения воды органическими веществами.
Криптомонады обитают в качестве эндосимбионтов в миксотрофных инфузориях.
Кроме того, криптомонады очень чувствительны к изменению окружающих условий, таких, как изменение рН, осмотических условий или температуры. Они в массе теряют трихоцисты, что приводит к быстрому разрушению клетки. С этим явлением связана значительная трудность по переводу криптомонад на лабораторные среды.
☼ Отдел ДИАТОМОВЫЕ
BACILLARIOPHYTA (DIATOMOPHYTA)
Диатомовые водоросли, или диатомеи – группа водорослей, отличающаяся наличием у клеток своеобразного «панциря», состоящего из кремнезёма (SiO2). Створки диатомовых водорослей не растворяются в большинстве природных вод, поэтому они осаждаются на протяжении последних 150 млн. лет, начиная с раннего мелового периода. Таким образом, есть основания полагать, что диатомеи появились до наступления мелового периода.
Численность: более 10 тысяч видов.
Среда обитания: диатомовые водоросли распространены повсеместно. Они обитают в соленых и пресных водоемах, на влажной почве, скалах, корке стволов деревьев и т. д. Много их в иле, на дне водоемов.
Уровень организации: к отделу относятся микроскопические одноклеточные или колониальные по организации водоросли, лишенные подвижных жгутиковых стадий (за исключением наиболее примитивных).
Образ жизни: диатомовые встречаются как в планктоне, так и в перифитоне, и бентосе. Диатомеи занимают господствующее положение среди обрастаний высших растений и макроскопических водорослей в пресных водоёмах и морях. Обрастанию могут подвергаться многие животные (такие водоросли называются эпизоонтами) от ракообразных до китов. Среди диатомей встречаются и эндобионты, которые обитают в других организмах, например, в бурых водорослях, фораминиферах.
Структура таллома: коккоидная.
Строение клетки. Характерная особенность диатомовых – в строении клеточной оболочки. К наружному уплотненному слою протопласта клетки плотно прилегает панцирь из кремниевого гидрогеля (рис. 70). Панцирь имеет две отдельные части (теки), плотно надетые друг на друга, как крышка на коробку: верхнюю – эпитеку и нижнюю – гипотеку. Каждая из этих частей состоит из створки («донышка») – плоской стороны с немного загнутыми краями, и пояска – узкого кольца, плотно соединенного со створкой. Поясок эпитеки плотно надвинут на поясок гипотеки. В створках имеются сквозные отверстия – поры, а также пустоты. Скульптура поверхности створок очень разнообразна и имеет систематическое значение.
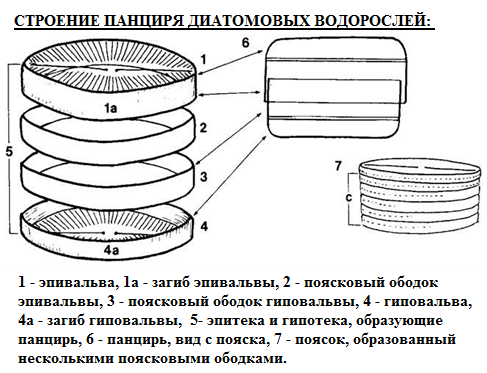
Рис. 70. Строение панциря диатомовых водорослей 1 – эпивальва, 1а – загиб эпивальвы, 2 – поясковый ободок эпивальвы, 3- поясковый ободок гиповальвы, 4 – гиповальва, 4а – загиб гиповальвы, 5 – гипотека и эпитека, образующие панцирь, 6 – панцирь, вид с пояска, 7 – поясок, образованный несколькими поясковыми ободками..
Большая часть клетки диатомовой водороли заполнена вакуолью, а цитоплазма располагается тонким слоем вдоль стенок. Ядро обычно расположено в центре клетки, в особом цитоплазменном мостике.
В хроматофорах содержатся хлорофиллы а и с, фукоксантин, придающий талломам буроватый цвет. Запасные вещества – масло, волютин, хризоламинарин.
Тип питания: автотрофы и миксотрофы. Некоторые способны переходить от автотрофного питания к гетеротрофному. Очень немногие формы утратили фотосинтетические пигменты и стали гетеротрофами.
Размножение происходит 1) вегетативным путем при помощи деления;
2) либо половым процессом в виде анизогамии, изогамии, реже оогамии (рис. 71).
Изо- и анизогамный процесс осуществляется посредством лишенных жгутиков гамет. При оогамном процессе мужская гамета имеет жгутик. Жгутик этот уникален тем, что не имеет центральных микротрубочек.
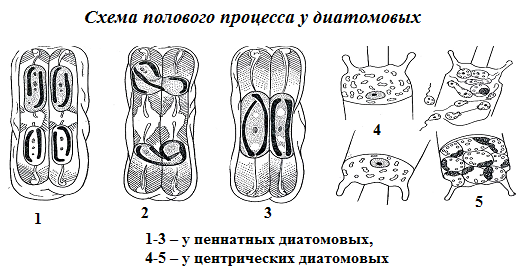
Рис. 71. Схема полового процесса у диатомовых: 1-3 – у пеннатных диатомовых; 4-5 – у центрических диатомовых.
Вегетативное размножение осуществляется митотическим делением протопласта. Затем дочерние протопласты расходятся, и каждый достраивает новую гипотеку. При этом оставшаяся от материнской клетки гипотека становится эпитекой дочерней клетки. Серия таких делений ведет к последовательному измельчению особей (рис. 72).

Рис. 72. Вегетативное размножение у диатомовых.
Восстановление исходных размеров клетки происходит в результате полового процесса, приводящего к образованию ауксоспор (растущих спор). Предположительно, образование ауксоспор связано с мельчанием клеток в результате их вегетативного размножения и необходимости восстановления размеров. Ауксоспорообразование всегда связано с половым процессом (рис. 73).
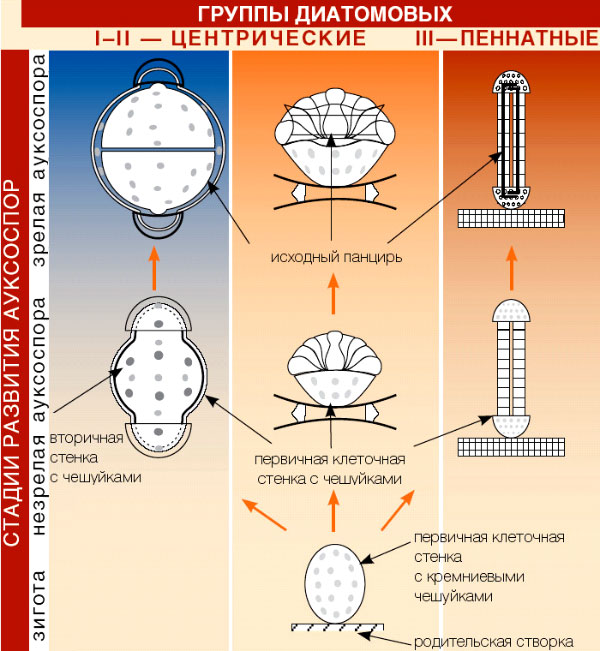
Рис. 73. Стадии развития ауксоспор у диатомовых.
У пеннатных диатомей сближаются две клетки, створки раздвигаются, происходит редукционное деление ядер, после чего гаплоидные ядра разных клеток попарно сливаются, и образуется одна или две аукоспоры (остальные гаплоидные ядра редуцируются). У центрических водорослей аукоспора образуется из одной клетки, в которой сначала происходит деление материнского диплоидного ядра на четыре гаплоидных ядра, два из них редуцируются, а два сливаются, образуется зигота, которая без стадии покоя резко увеличивается в размерах и образует ауксоспору. После созревания ауксоспоры в ней развивается новая клетка, у которой сначала формируется эпитека, затем гипотека, нормальных размеров. Все диатомовые водоросли – диплоидные организмы.
Жизненный цикл диатомовых водорослей проходит в диплоидной фазе, гаплоидны у них только гаметы, образующиеся непосредственно перед половым процессом.
Роль в природе, жизни человека и в биогеоценозах. Диатомовые в водных экосистемах доминируют над другими микроскопическими водорослями круглый год. Являясь важнейшей составляющей морского планктона, диатомовые создают до четверти всего органического вещества планеты.
Диатомовые водоросли играют большую роль в круговороте кремния в природе: они ежегодно извлекают из воды и используют для построения своих панцирей до 150 х109 т кремнезема, а отмирая, образуют особые геологические породы – диатомиты и трепел (рис. 74).

Рис. 74. Добыча диатомита.
Обладающие высокой пористостью и низким удельным весом, диатомиты широко используются в различных отраслях промышленности – их используют при производстве взрывчатки (динамита), материалов для звуковой и тепловой изоляции (кирпичи), при шлифовке металлов. Очень широкое применение нашли диатомиты в качестве фильтрующего материала при производстве различных масел, жиров, в сахарной и химической промышленности.
В медицине они используются для изготовления фильтров и адсорбционных поверхностей.
Систематика. Деление отдела на классы производится на основании строения панциря.