

Электрические явления в возбудимых тканях.
Электрическая сигнализация основана на выработке электрического заряда и последовательного его распространения. Разность потенциалов – это разность концентраций электрически заряженных частиц. Это следствие разделения в среде электрически заряженных частиц (φ – разность потенциалов). ЭДС обеспечивает движение – к + и наоборот и возникновение электрического тока.
В основе электрической сигнализации лежит изменение φ и ЭДС:
мембранный потенциал покоя (МПП)
мембранный потенциал действия (МПД)
рецепторный потенциал (РП)
пресинаптический и постсинаптический биопотенциалы (ПСП, ПоСП)
Потенциал покоя – относительно устойчивая разность потенциалов между внутренней и наружной поверхностью мембраны клетки, которая находится в состоянии физиологического покоя. Снаружи+, внутри–. Величина: -30 - -90 мВ. φ покоя рассчитан (теоретически) – 80-90 мВ. φ покоя реально φп для: нейрона – 60-(-70) мВ; скел мышечной клетки – 90 мВ.
Проницаемость в покое: Рк: РN0: Рсl=1:0,04:0,045
Возбуждение: Рк: РN0: Рсl = 1:20:0,45 РNa↑ в 500 раз.
Распределение ионов.
Миоцит |
Цитоплазма (мМ/L) |
Внекл жидкость (мМ/L) |
Na+ |
12 |
142 |
K+ |
140 |
4 |
Cl- |
4 |
120 |
Другие анионы. |
148 |
0 |
Нейрон |
Цитоплазма (мМ/L) |
Внекл жидкость (мМ/L) |
Na+ |
15 |
150 |
K+ |
150 |
55 |
Cl- |
9 |
125 |
Равновесный потенциал – это потенциал равновесия (или равенства скоростей) выхода или входа ионов одного вида.
Равновесный К+ потенциал
![]()
При
![]() ,
Ек=97,5 мВ, ЕNa=+55,
ЕCl=-0,85 мВ
,
Ек=97,5 мВ, ЕNa=+55,
ЕCl=-0,85 мВ
Плазматическая мембрана имеет φ, который является следствием:
1) работы Na-K насоса (повышает выход калия)
2) разная проницаемость для разных ионов (натрия и калия)
3) накопление молекул белка и органических соединений в примембранной области, имеющих отрицательный заряд.
4) ионы К+ под действием сил конц.град.легко диффундируют через каналы наружу, они концентрируются у внеш.поверх. мембраны, удерживаясь силами эл.поля отрицательных зарядов внутренней поверхности мембраны кл. В клетке возникает донановский φ (если мембраны “+” ионы пропускает, а “-” нет.)
5) Na+ не может пройти через мембрану внутрь клетки через ионные поры, т.к. хорошо ассоциируется с др в-вами, имеет гидратную оболочку и каналы для него закрыты.
МПП для каждого вида клеток достаточно постоянный. Это связано со строением клеточных мембран (ионных пор, каналов, плотностью расположения насосов).
Роль: 1. Регуляция проницаемости (глюкоза, амк). 2. Основа для возникновения и кодирования биопотенциалов.
Биопотенциал:
1. Носитель информации и субстрат ее передачи
2. Энергия БП используется для синтеза АТФ в клетках
3. Энергия БП используется для транспорта через мембрану:
а) пора б) ионный канал в) ионообменник (интегративный белок с двумя центрами) г) активный первичный транспорт (против градиента концентрации с использованием АТФ).
4. Энергия БП используется для межклеточных взаимодействий (агрегация и адгезия)
Рецепторный потенциал – изменение разности потенциалов на рецепторной мембране, во время воздействия на неё раздражителем.
В структуру мембраны входят каналы. Вероятность нахождения канала в закрытом или открытом состоянии является статистической функцией механической деформации и смещения той части мембраны, в которой располагается канал. Мех.деформация мембраны ведёт к открытию ионных каналов. Онако изменение проницаемости может быть связано с появлением мех.дислокаций и изменений в стр-ре мембраны. Na+ идёт в клетку и деполяризует мембрану. Выход К+ восстанавливает исходный потенциал. Восстановление распределения ионов осуществляет Na-К-насос.
1. Амплитуда РП пропорциональна логарифму изменения силы давления.
2. Длительность РП пропорциональна длительности воздействия раздражителя.
3. Вместе 1. и 2. = аналоговое кодирование.
4. Не сопровождается перезарядкой мембраны чувствительного окончания.
5. Распространяется по мембране нервного окончания на небольшие расстояния с затуханием.
6. Может порождать локальные токи.
Потенциал действия – кратковременное колебание разности потенциалов мембраны возбудимой клетки, сопровождающееся изменением знака заряда.
Разный ПД в разных клетках обусловлен различным содержанием ПЗИК
Нервные клетки МПП = -80-90 мВ, сердечные -80-90, но Na-K каналов меньше, основную роль играют медленные кальциевые каналы. ГМК – нет калиевых каналов. МПП = -45-60 мВ, имеют кальциевые каналы с различных порогом действия.
Механизм развития потенциала действия
Причина – РП (генераторный потенциал).
В потенциале действия выделяют:
1. Фаза деполяризации (0,3-0,5 мс). Деполяризация – процесс абсолютного уменьшения отрицательного заряда внутренней стороны плазматической мембраны. Критическая точка деполяризации (КТД) – величина мембранного потенциала, при которой открываются все Na+-ПЗИК. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. РП вызывает возникновение локальных круговых токов между точками на участках мембраны рецептора немиелинизированного окончания и 1 перехвата Ранвье. Эти токи протекают вдоль и поперек мембраны нервного волокна. В мембране перехвата Ранвье большая плотность Na+-ПЗИК и К+-ПЗИК. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД = 10 мВ) происходит лавинообразное открытие Na+-ПЗИК на 0,5 мс. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
2. Фаза быстрой и медленной реполяризации. В результате деполяризации мембраны происходит открытие К+-ПЗИК. Ионы К+ выходят из клетки по градиенту концентрации, что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется (следовая отрицательность).
3. Фаза гиперполяризации (следовый положительный потенциал) развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+/K+ помпы. Причины: инерционность закрывающихся каналов.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение. Пороговый потенциал – разность между МПП и критическим уровнем деполяризации.
Абсолютная рефрактерность (полная невозбудимость): все Na+ каналы открыты. Обуславливает одностороннее распространение нервного импульса.
Относительная рефрактерность: Na+ каналы реактивирутся.
Повышенная возбудимость (супернормальность, экзальтация): низкий порог возбуждения, когда реполяризация достигнет Ек.
Пониженная возбудимость (субнормальность): величина порога больше, т.к. разница потенциалов увеличивается.
Фазы: 1. деполяризация; 2. реполяризация; 3.гиперполяризация.
100 %
t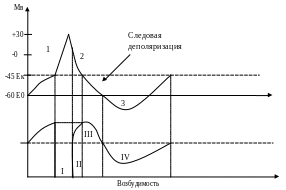
Лабильность - функциональная подвижность, скорость протекания элементарных физиологических процессов в клетке. Количественной мерой является max частота циклов возбуждения, которую может воспроизводить клетка. Чем короче рефрактерный период, тем больше лабильность клетки.
Сравнительная характеристика ЛО, РП, ПД.
ПД |
РП, ЛО |
Закон “все или ничего” |
Закон порога силы и времени |
Распространяется без затухания |
РП - С затуханием. ЛО – Не распространяется |
Не суммируются. |
Все суммируются |
Причина - РП |
Причина – внешнее воздействие |
Сопровождается перезарядкой мембраны |
Не сопровождается перезарядкой мембраны чувствительного окончания. |
Дискретное кодирование – процесс преобразования информации о характере воздействия в частоту нервного импульса. |
Аналоговое кодирование – процесс преобразования информации о силе воздействия в амплитуду и о продолжительности воздействия в длительность РП |
Электротонический потенциал (ЭП) – сдвиг мембранного потенциала клетки, вызываемый действием постоянного электрического тока подпороговой силы. ЭП есть пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов при этом не изменяется. ЭП не проявляется физиологической реакцией клетки, поэтому ЭП не является возбуждением.
Локальный ответ (ЛО) – активная реакция клетки на электрический раздражитель более сильного но подпорогового. Состояние ионных каналов и транспорт ионов при этом изменяется незначительно. ЛО не проявляется заметной физиологической реакцией клетки. ЛО называют местным возбуждением, т.к. это возбуждение не распространяется по мембранам возбудимых клеток.
Нервное волокно – отростки нервных клеток (аксоны и дендриты). Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
обладают высокой лабильностью, т.е. могут воспроизводить ПД с очень высокой частотой.
Миелиновые и безмиелиновые.
Афферентные и эфферентные
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты (шванновские клетки) сначала прикасаются к аксону, а затем окутывают его. В ЦНС – олигодедроциты. Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку). Миелиновая оболочка не является непрерывной – перехваты Ранвье через 0,5-2мм. В их области высокая плотность Na и К ПЗИК. Миелин – многослойная мембрана из ФЛ, ХС, белка и немного др.вв.
1. Аα волокна: 12-20 мкм, скорость 70-120м/с. Эфферентные от α-мотонейронов к экстрафузальным мыш.волокнам для инициации движения. Аγ волокна: 4-8мкм, 15-40м/с. Эфферентные от γ-мотонейронов к мышечным веретёнам для координации движения и афферентные от рецепторов прикосновения. Аβ волокно: 8-12 мкм, 40-70м/с. Афферентные от рецепторов прикосновения. Аδ волокно: 1-4 мкм, 5-15 м/с. Афферентные от рецепторв тепла, боли, давления.
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В 1 леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки.
2. В волокно: 1-3,5 мкм, 3-18 м/с. Преганглионарные вегетативные.
3. С волокно: 0,5-2 мкм, 0,5-3 м/с. Постганглионарные вегетативные, афферентные от рецепторов тепла, боли, давления.
Механизм проведения возбуждения по безмиелиновым нервным волокнам.
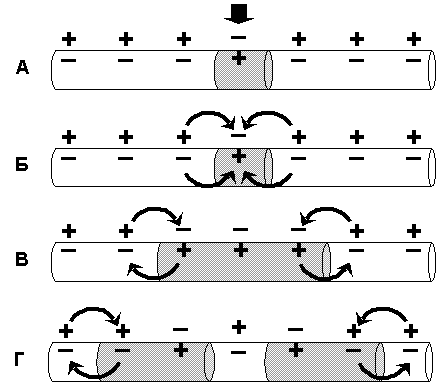
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет - заряд, а наружная сторона мембраны – +. Во время развития ПД в возбужденном участке мембраны происходит реверсия заряда (рис. 4.2 А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 4.2 Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 4.2 В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 4.2 Г). Таким образом, волна возбуждение охватывает все новые участки мембраны нервного волокна.
Механизм проведения возбуждения по миелиновым нервным волокнам.
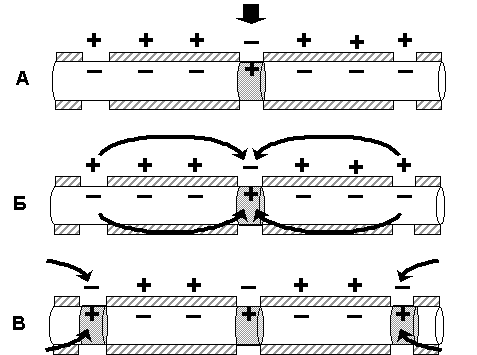
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только участках мембраны, расположенных в области перехватов Ранвье. При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис 4.3 А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис 4.3 Б). Однако, в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис 4.3 В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Законы проведения возбуждения по нервным волокнам.
Закон анатомической и физиологической непрерывности - возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В афферентных возбуждение распространяется от 1 перехвата к телу клетки. В эфферентных: от аксонного холмика к окончанию.
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна. Из-за более высокого эл.сопротивления между разными нервными стволами, по сравнению с соседними участками одного нервного ствола.
Аксонный транспорт.
Антеградный – от тела к окончаниям.
Быстрый 25-40 см/сут. Обеспечитвает перемещение медиаторов, некоторых органелл и ферментов. С помощью АТФ, актина, Са2+ и цитоскелета.
Медленный 1-2 мм/сут. Соответствует скорости роста акона при регенирации. Движение аксоплазмы с органеллами, РНК, белками, БАВ.
Ретроградный – от окончания к телу. 30 см/сут. Транспорт АХэстеразы, разруш.органелл, в-в, регулирующих синтез белка, вирусов, токсинов.
Синапс – специализированная структура, обеспечивающая межклеточную передачу сигналов эл./хим. природы.
Классификация:
Электрические – между кл., формируются плотные щелевые контакты. 3 нм. Общие ионные каналы, через которые с помощью ионных токов осущ.передача. Эмбриональный мозг, глия, ГМЦ, КМЦ, железистые кл. Постсинаптический потенциал 1 мВ и могут вызвать ПД.
Химические – специализ.стр-ры в области контакта. Щель 20-50нм. Центральные и переферические.
Нейромедиаторы:
1. Амины: холинэргические – АЦХ, адренэргические – А и НА, серотонинэргические – серотонин, допаминэргические – допамин.
Аминокислоты: Глу, Асп, ГАМК, Гли.
Пептиды: в-во П, ВИП, соматостатин, динорфин, энкефалин, нейропептид Y, холецистокинин, тиреолиберин.
2. Возбуждающие: Глу, Асп. Тормозные: ГАМК, Гли.
3. Нейромедиаторы – вызывают изменения на постсинаптической мембране менеее чем за 1 мс и длится он менее 10 мс. Нейромодуляторы – более продолжительное действие и способны регулировать действие нейромедиатора.
Функциональные св-ва синапсов:
Одностороннее проведение сигнала
Замедленная передача сигнала 0,5-2мс
Способность к временной суммации эффекта (2 сигнала подряд за 1-10мс)
Трансформация ритма возбуждения (частота импульсов, приходящих к пресинаптической мембране не равна частоте ПД, искл. – с нерва на мышцу.)
Низкая лабильность и высокая утомляемость (50-100имп/с, в 5-10 раз меньше, чем у нерв.вол.)
Высокочувствительны к БАВ, лекарствам, ядам (стрихнин – связ. рецепторы к Гли, столбнячный токсин – нарушает выделение медиатора)
Облегчение синаптической передачи (если импульсы с частотой 10-50 в мин)
Депрессия синаптической передачи (если импульсы очень частые, более 100Гц).
Структура: Нейромедиатор – везикулы 50 нм, синтез в теле нейрона. Неромодуляторы – везикулы 100 нм, синтез в шЭПР и АГ. В пресинптической терминали: синаптотагмин, синаптобревин, синапсин, ассоциированы с везикулами и связывают их с цитоскелетом. 1. Деполяризация пресинптической терминали (РП например) – открытие Са2+ПЗИК (внутри его 10-8, снаружи 10-4) – слияние везикул (синаптотагмин) – мембраны везикул используются повторно. Нейропептиды высвобождаются медленнее и при более высоких концентрациях Са, т.е. при высокой частоте импульсов. Са2+насос выкачивает Са2+. В адренэргических синапсах на пресинп.мембране есть ауторецепторы, через которые осущ.реаптейк нейромедиатора из щели.
2. В синп.щели медиатор диффундирует по град.конц. к постсинптич.мембр., где связ. со своими рецепторами. Тут же нейромедиатор разрушается холинэстеразой.
3. Постсинптич.мембрана: а) лигандзависимый ионный канал (ионотропный рецептор) – генерация ПСП. б) 7-ТМС (метаботропный рецептор) – ПСП и изменение метаболизма.
ВПСП
– (АЦК с н-Хр, Глу с АМРА-р) – открытие неселективного катионного канала – вход Na+ (по град.заряда и по град.конц.) опережает выход К+ (по град.конц.) - кратковременное колебание разности потенциалов = ВПСП.
Может распространяться на небольшие расстояния с затуханием, порождая локальные токи, не сопровождается сменой знака заряда.
– (Глу с NMDA-р) – открытие неселективного катионного – вход Na+, Са2+ опережают выход К+ = ВПСП и Са2+ изменяет метаболизм, экспрессию генов, как вторичный посредник.
ТПСП
– (ГАМК, Гли) – открытие лигандзависимого Cl- канала – вход Cl- – гиперполяризация – ТПСП.
Может распространяться на небольшие расстояния с затуханием, порождая локальные токи (противоположны ВПСП), которые вызывают гиперполяризацию.
Нейро-мышечный синапс. Образован утолщенным окончанием аксона моторного нейрона (бляшка) и мышечным волокном поперечно-полосатой мускулатуры (часть сарколеммы).
Пресинап.мембрана: много митохондрий (Ацетил-КоА и АТФ), ацетилхолинтрансфераза. Большая протяженность, чем в ЦНС.
Синаптическая щель: 50-100 нм, ацетилхолинэстераза.
Концевая пластинка: много впячиваний, 20млн н-ХРмыш.типа – неселективные Na и К каналы.
Нервный импульс – деполяризация мембраны пресинп.терминали.
Открытие ПЗ(Ca2+)К – вход С2+
Высвоюождение АЦХ
Диффузия АЦХ и связывание с н-ХРмт
Открытие неселективных ЛЗ(Na+иК+)К - вход Na+ и выход К+
Деполяризация (Потенциал Концевой Пластинки = ВПСП)
Возник. Лок. круговых токов между конц. пласт. и ближайшим участком сарколеммы
Деполяризация сарколеммы, если Ек, то ПД.
Концевая пластинка не может генерировать ПД, т.к. не имеет ПЗ(Na+)К, которые обеспечивают быстрый вход Na+ и смену знака заряда мембраны.
9. Когда конц.АЦХ снизится до 10 нмолей АЦХ освобождает рецепторы.
Синаптическая задержка = 1 мс.
Миниатюрные потенциалы концевой пластинки – в покое 1 пузырёк в секунду выходит из пресинап.мембраны. Вызывает очень малую деполяризацию – 0,1-0,2мВ. Трофическое влияние: стимулирует синтез каналообразующих белков, регулирует метаболизм, влияет на поддержание тканевой специфичности.