

9. Матричный биосинтез белков. Транскрипция.
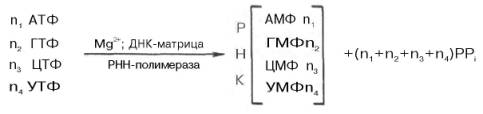
транскрипция, несмотря на кажущуюся схожесть с репликацией, в частности химическим механизмом, направлением синтеза и использованием матрицы, отличается рядом особенностей: не требует синтеза праймера, использует не всю молекулу ДНК, а только ее отдельные короткие сегменты (отдельные гены или группы генов) и, наконец, требует наличия только одной из цепей ДНК в качестве матрицы, которая полностью сохраняется (при репликации ДНК она сохраняется наполовину). Геном каждой клетки человека состоит из 3,5•109 пар оснований; они могут обеспечить кодирование более 1,5•106 пар генов. Однако имеющиеся данные о количестве и разнообразии белков в организме человека (около 100000) свидетельствуют о том, что значительная часть генома человека не транскрибируется и соответственно не переводится на аминокислотную последовательность белков. Известно также, что определенная часть нетранслируемого генома человека выполняет регуляторную функцию в процессе экспрессии генов. Вмолекуле ДНК различают, кроме уникальных неповторяющихся последовательностей, содержащих кодирующие гены, также множество повторяющихся последовательностей (повторы), биологический смысл которых до сих пор неясен. Современные представления о механизме синтеза РНК в клетках в значительной степени обязаны открытию в 1960 г. в двух лабораториях США (Дж. Хервиц и С. Вейс) особого фермента – РНК-полимеразы, катализирующей синтез РНК из свободных нуклеозидтрифосфатов. Фермент требует наличия ионов Mg2+или Мn2+ и одновременного присутствия всех 4 типов рибонуклеозидтрифосфатов (АТФ, ГТФ, ЦТФ и УТФ). Самым удивительным свойством фермента оказалось то, что для включения нуклеотидов в РНК необходимо обязательное присутствие предобразо-ванной ДНК-матрицы . При тщательном изучении механизма синтезаРНК при участии РНК-полимеразы, называемой также ДНК-зависимой РНК-полимеразой (транскриптазой), было установлено, что молекула предобразованной ДНК, необходимая для реакции полимеризации, полностью определяет последовательность рибонуклеотидов во вновь синтезированной молекуле РНК. Другими словами, на матрице ДНК комплементарно строится полирибонуклеотид, являющийся копией первичной структуры ДНК, с той только разницей, что вместо тимидилового нуклео-тида ДНК в РНКвключается уридиловый нуклеотид. Реакция синтеза РНК в общем виде может быть представлена следующим образом:
 В
синтезируемой молекуле РНК отдельные
мононуклеотиды, как и в ДНК,
связаны между собой 3'-5'-фосфодиэфирными
мостиками. Кроме того, сам механизм
действия фермента РНК-полимеразы во
многом совпадает с таковыми ДНК-полимеразы:
синтез также идет в направлении 5'–>3',
цепь РНК имеет
полярность, противоположную цепи
предобразованной ДНК.
Однако выявлены и существенные
различия. РНК-полимераза Е.
coli предпочтительнее функционирует в
присутствии нативной двухцепочечной
ДНК;
в опытах in vitro обе цепи ДНК копируются РНК-полимеразой;
in vivo транскрибируется, вероятнее всего,
только одна цепь ДНК.
Предполагают, что РНК-полимеразасвязывается
с одной цепью нативной ДНК в
определенной точке, вызывая расплетение
биспиральной структуры на ограниченном
участке, где и происходит синтез РНК.
Данные свидетельствуют, что у E. coli,
скорее всего, имеется единственная
ДНК-зави-
В
синтезируемой молекуле РНК отдельные
мононуклеотиды, как и в ДНК,
связаны между собой 3'-5'-фосфодиэфирными
мостиками. Кроме того, сам механизм
действия фермента РНК-полимеразы во
многом совпадает с таковыми ДНК-полимеразы:
синтез также идет в направлении 5'–>3',
цепь РНК имеет
полярность, противоположную цепи
предобразованной ДНК.
Однако выявлены и существенные
различия. РНК-полимераза Е.
coli предпочтительнее функционирует в
присутствии нативной двухцепочечной
ДНК;
в опытах in vitro обе цепи ДНК копируются РНК-полимеразой;
in vivo транскрибируется, вероятнее всего,
только одна цепь ДНК.
Предполагают, что РНК-полимеразасвязывается
с одной цепью нативной ДНК в
определенной точке, вызывая расплетение
биспиральной структуры на ограниченном
участке, где и происходит синтез РНК.
Данные свидетельствуют, что у E. coli,
скорее всего, имеется единственная
ДНК-зави-
симая РНК-полимераза, которая катализирует синтез всех типов клеточных РНК.