

Экзамен по биологии. I курс.
Раздел II
№1
Биология – наука о закономерностях и механизмах жизнедеятельности
Биология — наука о жизни, которая изучает закономерности жизни и развития живых существ. Термин «биология» был предложен немецким ботаником Г.Р. Тревиранусом и французским естествоиспытателем Ж.-Б. Ламарком в 1802 году независимо друг от друга.
Биология относится к естественным наукам. Разделы науки биологии можно классифицировать по-разному. Например, в биологии выделяют науки по объектам исследования: о животных — зоологию; о растениях — ботанику; анатомию и физиологию человека как основу медицинской науки. В пределах каждой из этих наук имеются более узкие дисциплины. Например, в зоологии выделяют протозоологию, энтомологию, гельминтологию и другие.
Основные методы биологии:
Описательный – сбор и описание фактического материала.
Сравнительный – позволяет путём сопоставления выделить сходства и различия между организмами. Является основой систематики.
Исторический – связан с именем Дарвина (выявляет пути развития организмов).
Экспериментальный – постановка опытов и изменение течения эксперимента в нужных опытах.
№2
Критерии и уровни организации живых организмов
К основным свойствам живого можно отнести:
Химический состав. Живые существа состоят из тех же химических элементов, что и неживые, но в организмах есть молекулы веществ, характерных только для живого (нуклеиновые кислоты, белки, липиды).
Дискретность и целостность. Любая биологическая система (клетка, организм, вид и т.д.) состоит из отдельных частей, т.е. дискретна. Взаимодействие этих частей образует целостную систему (например, в состав организма входят отдельные органы, связанные структурно и функционально в единое целое).
Структурная организация. Живые системы способны создавать порядок из хаотичного движения молекул, образуя определенные структуры. Для живого характерна упорядоченность в пространстве и времени. Это комплекс сложных саморегулирующихся процессов обмена веществ, протекающих в строго определенном порядке, направленном на поддержание постоянства внутренней среды — гомеостаза.
Обмен веществ и энергии. Живые организмы — открытые системы, совершающие постоянный обмен веществом и энергией с окружающей средой. При изменении условий среды происходит саморегуляция жизненных процессов по принципу обратной связи, направленная на восстановление постоянства внутренней среды — гомеостаза. Например, продукты жизнедеятельности могут оказывать сильное и строго специфическое тормозящее воздействие на те ферменты, которые составили начальное звено в длинной цепи реакций.
Самовоспроизведение. Самообновление. Время существования любой биологической системы ограничено. Для поддержания жизни происходит процесс самовоспроизведения, связанный с образованием новых молекул и структур, несущих генетическую информацию, находящуюся в молекулах ДНК.
Наследственность. Молекула ДНК способна хранить, передавать наследственную информацию, благодаря матричному принципу репликации, обеспечивая материальную преемственность между поколениями.
Изменчивость. При передаче наследственной информации иногда возникают различные отклонения, приводящие к изменению признаков и свойств у потомков. Если эти изменения благоприятствуют жизни, они могут закрепиться отбором.
Рост и развитие. Организмы наследуют определенную генетическую информацию о возможности развития тех или иных признаков. Реализация информации происходит во время индивидуального развития — онтогенеза. На определенном этапе онтогенеза осуществляется рост организма, связанный с репродукцией молекул, клеток и других биологических структур. Рост сопровождается развитием.
Раздражимость и движение. Все живое избирательно реагирует на внешние воздействия специфическими реакциями благодаря свойству раздражимости. Организмы отвечают на воздействие движением. Проявление формы движения зависит от структуры организма.
Выделяют уровни организации всего живого:
Молекулярно-генетический – это макромолекулы белков, полисахаридов, нуклеиновых кислот и гены. Явления уровня: пластический и энергетический обмен, передача наследственной информации.
Субклеточный – органоиды клетки. Явления уровня: функционирование органоидов.
Клеточный.
Тканевый. Единица уровня – группа клеток, выполняющая одинаковые функции. Явления уровня: дифференцировка клеток.
Организменный – единица будет являться особь в стадии онтогенеза. Явления уровня: те изменения, которые происходят с особью в процессе онтогенеза.
Популяционно-видовой. Единица уровня – популяция. Явления уровня: элементарные эволюционные преобразования.
Биогеоценотический – единицы уровня – биогеоценоз. Явления уровня: круговороты различных веществ.
Биосферный.
№3
Клеточная теория
Клетка - основная структурная, функциональная и генетическая единица организации живого, элементарная живая система. Клетка может существовать как отдельный организм (бактерии, простейшие, некоторые водоросли и грибы) или в составе тканей многоклеточных животных, растений, грибов. Термин «клетка» был предложен английским исследователем Робертом Гуком в 1665 г. Впервые используя микроскоп для изучения срезов пробки, он заметил множество мелких образований, похожих на ячейки пчелиных сот. Роберт Гук дал им название ячейки или клетки. Работы Р. Гука вызвали интерес к дальнейшим микроскопическим исследованиям организмов. Возможности светового микроскопа в XVII-XVIII веках были ограничены. Накопление материала о клеточном строении растений и животных, о структуре самих клеток шло медленно. Только в тридцатых годах XIX века были сделаны фундаментальные обобщения о клеточной организации живого.
Основные положения клеточной теории:
Клетка – элементарная единица всего живого
Все клетки разных организмов гомогенны по своему строению и свойствам.
Клетка – единая система, включающая в себя множества взаимосвязанных элементов, представляющих собой целостное образование.
Клетки увеличиваются в числе путём деления исходной клетки после удвоения её генетического материала (введено в 1959 году немецким патологоанатомом Рудольфом Вирховом: «Клетка от клетки»).
Многоклеточный организм - это сложный ансамбль объединённых и интегрированных в системы органов и тканей, связанных с друг другом с помощью физических, гуморальных и нервных факторов.
Клетки тотипотентны – то есть обладают одинаковым генетическим потенциалом и равноценны по генетической информации, но различной экспрессией (работой) генов, что приводит к морфологическому и функциональному разнообразию.
Клетка – это ограниченная активной мембраной упорядоченная системой биополимеров, участвующих в единой совокупности метаболических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
№4
Современные представления об организации эукариотической клетки
Основная часть поверхностного аппарата клетки - плазматическая мембрана. Клеточные мембраны — важнейший компонент живого содержимого клетки построены по общему принципу. Согласно жидкостно-мозаичной модели, предложенной в 1972 г. Николсоном и Сингером, в состав мембран входит бимолекулярный слой липидов, в который включены молекулы белков (рис. 15). Липиды — это водонерастворимые вещества, молекулы которых имеют два полюса, или два конца. Один конец молекулы обладает гидрофильными свойствами, его называют полярным. Другой полюс гидрофобный, или неполярный. В биологической мембране молекулы липидов двух параллельных слоев обращены друг к другу неполярными концами, а их полярные полюса остаются снаружи, образуя гидрофильные поверхности. Кроме липидов, в состав мембраны входят белки. Их можно разделить на три группы: периферические, погруженные (полуинтегральные) и пронизывающие (интегральные). Большинство белков мембраны является ферментами. Полуинтегральные белки образуют на мембране биохимический «конвейер», на котором в определенной последовательности осуществляется превращение веществ. Положение погруженных белков в мембране стабилизируется периферическими белками. Интегральные белки обеспечивают передачу информации в двух направлениях: через мембрану в сторону клетки и обратно. Интегральные белки бывают двух типов: переносчики и каналообразующие. Последние выстилают пору, заполненную водой. Через нее осуществляется прохождение ряда растворенных неорганических веществ с одной стороны мембраны на другую.
Гиалоплазма (основная плазма, матрикс цитоплазмы или цитозоль) основное вещество цитоплазмы, заполняющее пространство между клеточными органеллами. Гиалоплазма содержит около 90% воды и различные белки, аминокислоты, нуклеотиды, жирные кислоты, ионы неорганических соединений, другие вещества. Крупные молекулы белка образуют коллоидный раствор, который может переходить из золя (невязкое состояние) в гель ( вязкий). В гиалоплазме протекают ферментативные реакции, метаболические процессы ( гликолиз), синтез аминокислот, жирных кислот. На рибосомах, свободно лежащих в цитоплазме, происходит синтез белков. Гиалоплазма содержит множество белковых филаментов ( нитей), пронизывающих цитоплазму и образующих цитоскелет. В клетках животных организатором цитоскелета является область, расположенная рядом с ядром, содержащая пору центриолей (рис. 25, 26). Цитоскелет определяет форму клеток, обеспечивает движение цитоплазмы, называемое циклозом.
Эндоплазматическая сеть (ЭПС) - это система цистерн и каналов, «стенка» которых образована мембраной. ЭПС пронизывает цитоплазму в разных направлениях и делит ее на изолированные отсеки (компартменты). Благодаря этому в клетке осуществляются специфические биохимические реакции. Эндоплазматическая сеть выполняет также синтетическую и транспортную функции. Если на поверхности эндоплазматической мембраны есть рибосомы, ее называют шероховатой, если рибосом нет – гладкой (рис. 27). На рибосомах осуществляется синтез белков. Белки проходят через мембрану в цистерны ЭПС, где приобретают третичную структуру и транспортируются по каналам к месту потребления. На гладкой ЭПС происходит синтез липидов, стероидов. ЭПС — основное место биосинтеза и построения мембран цитоплазмы. Отчленяющиеся от нее пузырьки представляют исходный материал для других одномембранных органелл: аппарата Гольджи, лизосом, вакуолей.
Аппарат Гольджи - органелла, обнаруженная в клетке итальянским исследователем Камилло Гольджи в 1898 г. Аппарат Гольджи обычно расположен около клеточного ядра. Наиболее крупные аппараты Гольджи находятся в секреторных клетках. Основным элементом органеллы является мембрана, образующая уплощенные цистерны - диски. Они располагаются друг над другом. Каждая стопка Гольджи (у растений называемая диктиосомой) содержит от четырех до шести цистерн. Края цистерн переходят в трубочки, от которых отчленяются пузырьки (пузырьки Гольджи), транспортирующие заключенное в них вещество к месту его потребления. Отчленение пузырьков Гольджи происходит на одном из полюсов аппарата. Со временем это приводит к исчезновению цистерны. На противоположном полюсе аппарата осуществляется сборка новых дисков-цистерн. Они формируются из пузырьков, отпочковывающихся от гладкой эндоплазматической сети. Содержимое этих пузырьков, «унаследованное» от ЭПС, становится содержимым аппарата Гольджи, в котором подвергается дальнейшей переработке. Функции аппарата Гольджи разнообразны: секреторная, синтетическая, строительная, накопительная. Одна из важнейших функций - секреторная. В цистернах аппарата Гольджи происходит синтез сложных углеводов (полисахаридов), осуществляется их взаимосвязь с белками, приводящая к образованию мукопротеидов. С помощью пузырьков Гольджи готовые секреты выводятся за пределы клетки. Аппарат Гольджи образует гликопротеин (муцин), представляющий важную составную часть слизи; участвует в секреции воска, растительного клея. Иногда аппарат Гольджи принимает участие в транспорте липидов. В аппарате Гольджи происходит укрупнение белковых молекул. Он участвует в построении плазматической мембраны и мембран вакуолей. В нем формируются лизосомы.
Лизосомы (от греч. лизис – разрушение, расщепление, сома – тело) - пузырьки больших или меньших размеров, заполненные гидролитическими ферментами (протеазами, нуклеазами, липазами и другими). Лизосомы в клетках не представляют собой самостоятельных структур. Они образуются за счет активности эндоплазматической сети и аппарата Гольджи и напоминают секреторные вакуоли. Основная функция лизосом - внутриклеточное расщепление и переваривание веществ, поступивших в клетку или находящихся в ней, и удаление из клетки. Выделяют первичные и вторичные лизосомы (пищеварительные вакуоли, аутолизосомы, остаточные тельца). Первичные лизосомы представляют собой пузырьки, ограниченные от цитоплазмы одинарной мембраной. Ферменты, находящиеся в лизосомах, синтезируются на шероховатой эндоплазматической сети и транспортируются к аппарату Гольджи. В цистернах аппарата Гольджи вещества подвергаются дальнейшим превращениям. Пузырьки с набором ферментов, отделившиеся от цистерн аппарата Гольджи, называют первичными лизосомами (рис. 31). Они участвуют во внутриклеточном пищеварении и иногда секреции ферментов, выделяющихся из клетки наружу. Это происходит, например, при замене хряща костной тканью в процессе развития, при перестройке костной ткани в ответ на повреждение. Секретируя гидролитические ферменты, остеокласты (клетки-разрушители) обеспечивают разрушение минеральной основы и органического остова матрикса кости. Накапливающиеся «обломки» подвергаются внутриклеточному перевариванию. Остеобласты (клетки-строители) создают новые элементы кости. Первичные лизосомы могут сливаться с фагоцитарными и пиноцитарными вакуолями, образуя вторичные лизосомы. В них происходит переваривание веществ, поступивших в клетку путем эндоцитоза, усвоение их. Вторичные лизосомы— пищеварительные вакуоли, ферменты которых доставлены с помощью мелких первичных лизосом. Вторичные лизосомы (пищеварительные вакуоли) у простейших (амеб, инфузорий) - это способ поглощения пищи. Вторичные лизосомы могут выполнять защитную функцию, когда, например, лейкоциты (фагоциты) захватывают и переваривают попавшие в организм бактерии. Продукты переваривания поглощаются клеткой, но часть материала может остаться непереваренной. Вторичные лизосомы, содержащие нерасщепленный материал, называют остаточными тельцами или телолизосомами. Остаточные тельца обычно через плазматическую мембрану выводятся наружу (экзоцитоз). У человека при старении организма в остаточных тельцах клеток мозга, печени и в мышечных волокнах накапливается «пигмент старения» - липофусцин. Аутолизосомы (аутофагирующие вакуоли) присутствуют в клетках простейших, растений и животных. В этих лизосомах происходит разрушение отработанных органелл самой клетки (ЭПС, митохондрий, рибосом, гранул гликогена, включений и др.). Например, в клетках печени среднее время жизни одной митохондрии — около 10 дней. После этого срока мембраны эндоплазматической сети окружают митохондрию, образуя аутофагосому. Аутофагосомы сливаются с лизосомой, образуя аутофаголизосому, в которой происходит процесс распада митохондрии. Процесс уничтожения структур, ненужных клетке, называется аутофагией. Число аутолизосом возрастает при повреждениях клетки. В результате высвобождения содержимого лизосом в цитоплазму происходит саморазрушение клетки или аутолиз. При некоторых процессах дифференцировки аутолиз может быть нормой. Например, при исчезновении хвоста у головастика во время превращения его в лягушку. Ферменты лизосом принимают участие в аутолизе погибших клеток.
Известно более 25 генетических заболеваний, связанных с патологией лизосом. Например, в лизосомах может происходить накопление гликогена, если отсутствует соответствующий фермент.
№5
Современные представления об организации эукариотической клетки (Ядро)
Ядро было открыто и описано в 1833 г. англичанином Р. Броуном. Ядро присутствует во всех эукариотических клетках, за исключением зрелых эритроцитов и ситовидных трубок растений. Клетки, как правило, имеют одно ядро, но иногда встречаются многоядерные клетки. Ядро бывает шаровидной или овальной формы. В некоторых клетках встречаются сегментированные ядра. Размеры ядер - от 3 до 10 мкм в диаметре. Ядро необходимо для жизни клетки. Оно регулирует активность клетки. В ядре хранится наследственная информация, заключенная в ДНК. Эта информация, благодаря ядру, при делении клетки передается дочерним клеткам. Ядро определяет специфичность белков, синтезируемых в клетке. В ядре содержится множество белков, необходимых для обеспечения его функций. В ядре синтезируется РНК. Ядро имеет ядерную оболочку, отделяющую его от цитоплазмы, кариоплазму (ядерный сок), одно или несколько ядрышек, хроматин.
Ядерная оболочка состоит из двух мембран. В ней имеются поры, играющие важную роль в переносе веществ в цитоплазму и из нее. Поры не являются постоянными образованиями. Их число меняется в зависимости от функциональной активности ядра. Число пор увеличивается в период наибольшей ядерной активности. Ядерная оболочка связана непосредственно с эндоплазматической сетью. На наружной мембране ядерной оболочки, с внешней стороны находятся рибосомы, синтезирующие специфические белки, образующиеся только на рибосомах ядерной оболочки. Ядерный сок (кариоплазма) - внутреннее содержимое ядра, представляет собой раствор белков, нуклеотидов, ионов, более вязкий, чем гиалоплазма. В нем присутствуют также фибриллярные белки. В кариоплазме находятся ядрышки и хроматин. Ядерный сок обеспечивает нормальное функционирование генетического материала. Ядрышки - обязательный компонент ядра, обнаруживаются в интерфазных ядрах и представляют собой мелкие тельца, шаровидной формы. Ядрышки имеют большую плотность, чем ядро. В ядрышках происходит синтез р–РНК, других видов РНК и образование субъединиц рибосом. Возникновение ядрышек связано с определенными зонами хромосом, называемыми ядрышковыми организаторами. Число ядрышек определяется числом ядрышковых организаторов. В них содержатся гены р–РНК. Хроматин (окрашенный материал) – плотное вещество ядра, хорошо окрашиваемое основными красителями. В состав хроматина входят молекулы ДНК в комплексе с белками (гистонами и негистонами), РНК. В неделящихся (интерфазных) ядрах хроматин может равномерно заполнять объем ядра, находясь в деконденсированном состоянии. Этот диффузный хроматин (эухроматин) генетически активен. Молекулы ДНК, содержащие наследственную информацию, способны удваиваться при репликации, и возможна передача (транскрипция) генетической информации с ДНК на и–РНК. Иногда в интерфазном ядре бывают видны глыбки хроматина, представляющие собой участки конденсированного хроматина (гетерохроматина). Это неактивные участки. Например, в клетках женского организма, где присутствуют две X–хромосомы, одна находится в активном диффузном состоянии, а вторая в неактивном, конденсированном состоянии. Во время деления ядра хроматин окрашивается интенсивнее, происходит его конденсация – образование более спирализованных (скрученных) нитей, называемых хромосомами. Хромосомы синтетически неактивны. Строение хромосом лучше всего изучать в момент их наибольшей конденсации, т.е. в метафазе и начале анафазы митоза. Каждая хромосома в метафазе митоза состоит из двух хроматид, образовавшихся в результате редупликации, и соединенных центромерой (первичной перетяжкой). В центральной части центромеры находятся кинетохоры, к которым во время митоза прикрепляются микротрубочки нитей веретена. В анафазе хроматиды отделены друг от друга. Из них образуются дочерние хромосомы, содержащие одинаковую генетическую информацию. Центромера делит хромосому на два плеча. Хромосомы с равными плечами называют равноплечими или метацентрическими, с плечами неодинаковой длины - неравноплечими субметацентрическими, с одним коротким и вторым почти незаметным палочковидными или акроцентрическими. Некоторые хромосомы имеют вторичную перетяжку, отделяющую спутник. Вторичные перетяжки называют ядрышковыми организаторами. В них в интерфазе происходит образование ядрышка. В ядрышковых организаторах находится ДНК, отвечающая за синтез р-РНК. Плечи хромосом оканчиваются участками, называемыми теломерами, не способными соединяться с другими хромосомами.
Число, размер и форма хромосом в наборе у разных видов могут варьировать. Совокупность признаков хромосомного набора называют кариотипом.
Хромосомный набор специфичен и постоянен для особей каждого вида. У человека 46 хромосом, у мыши - 40 хромосом и т.д. В соматических клетках, имеющих диплоидный набор хромосом, хромосомы парные. Их называют гомологичными. Одна хромосома в паре происходит от материнского организма, другая - от отцовского. Изменения в структуре хромосом или в их числе возникают в результате мутаций. Каждая пара хромосом в наборе индивидуальна. Хромосомы из разных пар называют негомологичными. В кариотипе различают половые хромосомы (у человека это Х–хромосома и Y–хромосома) и аутосомы (все остальные). Половые клетки имеют гаплоидный набор хромосом. Основу хромосомы составляет молекула ДНК, связанная с белками (гистонами и др.) в нуклеопротеид. Основное положение молекулярной биологии, сформулированное Ф. Криком, утверждает, что перенос генетической информации осуществляется:
от ДНК к ДНК путем репликации
от ДНК через и-РНК (м-РНК) к белку
Процесс самовоспроизведения макромолекул нуклеиновых кислот (репликация) обеспечивает точное копирование генетической информации и передачу ее от поколения к поколению. Принцип комплементарности, лежащий в основе структуры молекулы ДНК, дает возможность понять, как синтезируются новые молекулы в синтетическом периоде интерфазы жизненного цикла клетки перед ее делением.
№6
Современные представления об организации эукариотической клетки (2хмембранные органеллы)
Митохондрии имеются во всех эукариотических клетках
Основная функция митохондрии связана с окислением органических соединений и использованием энергии, освобождающейся при распаде этих соединений, для синтеза молекул АТФ (рис. 35). Число, размеры, форма митохондрии в клетке различны и непостоянны. Митохондрии могут иметь вытянутую, округлую, спиральную, палочковидную форму. В клетках, нуждающихся в большом количестве энергии, митохондрии много. Например, в одной печеночной клетке их может быть около 1000. Локализация митохондрии различна. Обычно они скапливаются вблизи тех участков цитоплазмы, где велика потребность в энергии АТФ. Например, в скелетных мышцах митохондрии находятся вблизи миофибрилл. Каждая митохондрия окружена двумя мембранами. Наружная митохондриальная мембрана, отделяющая ее от гиалоплазмы, гладкая. Наружную мембрану от внутренней отделяет межмембранное пространство. Внутренняя мембрана, ограничивающая матрикс митохондрии, образует многочисленные складки (кристы). Чем больше крист присутствует в митохондрии, тем интенсивнее протекают окислительно-восстановительные процессы.
Например, митохондрии клеток сердечной мышцы содержат втрое больше крист, чем митохондрии клеток печени. В матриксе митохондрии находятся различные ферменты, кольцевая молекула ДНК, рибосомы, РНК. На митохондриальных рибосомах синтезируются белки, специфические для органеллы. Митохондрии относят к полуавтономным органеллам. На внутренней мембране присутствуют белки, катализирующие окислительновосстановительные реакции в дыхательной цепи, ферменты, участвующие в синтезе АТФ, и специфические транспортные белки. Наружная мембрана содержит ферменты, участвующие в синтезе митохондриальных липидов. Митохондрии называют энергетическими станциями клетки. В них происходит окисление органических веществ, благодаря чему освобождается заключенная в веществах энергия. Она необходима для осуществления всех жизненных процессов в клетке. Эта энергия используется на восстановительные процессы. В митохондриях осуществляется восстановление ( синтез) АТФ (аденозинтрифосфорной кислоты) из АДФ (аденозиндифосфорной кислоты). В результате энергия, выделившаяся при разложении веществ, вновь переходит в связанную форму в молекуле АТФ. АТФ транспортируется ко всем участкам клетки, где необходима энергия. Эта энергия, заключенная в макроэргических связях в молекуле АТФ, выделяется при распаде АТФ до АДФ. АДФ снова поступает в митохондрии, где в ходе восстановительных реакций превращается в АТФ, связав энергию, освобожденную при окислении веществ. Окислительно-восстановительные процессы в митохондриях протекают ступенчато, при участии окислительных ферментов. Эти процессы обусловлены переходом энергии химических связей, заключенной в веществах, в макроэргическую связь в молекуле АТФ, которая синтезируется при использовании освобождающейся энергии из АДФ и фосфата. Митохондрии размножаются поперечным делением или фрагментацией на более короткие.
Пластиды – двумембранные органеллы, присутствующие в растительных клетках. Различают три вида пластид: хлоропласты, хромопласты и лейкопласты (рис. 36). Хлоропласты – органеллы, осуществляющие фотосинтез, ограничены двумя мембранами – внешней и внутренней. Между мембранами есть межмембранное пространство. В хлоропластах присутствует зеленый пигмент – хлорофилл, находящийся в системе мембран, которые погружены во внутреннее содержимое пластид – матрикс (или строму). В строме хлоропластов находятся плоские мембранные структуры, называемые ламеллами. Ламеллы стромы лежат параллельно друг другу и связаны между собой. Две соседние мембраны, соединяясь концами, формируют замкнутые плоские мембранные структуры в форме диска – тилакоиды, – содержащие внутри жидкость. Тилакоиды, уложенные в стопки, образуют граны. Число тилакоидов на одну грану варьирует: от нескольких единиц до 50 и более. Тилакоиды в гране тесно сближены друг с другом. В состав граны, кроме замкнутых дисков тилакоидов, входят участки ламелл. Ламеллы стромы связывают между собой отдельные граны хлоропласта. Количество гран в хлоропластах может достигать 40-60. В мембранных структурах хлоропластов присутствуют пигменты: зеленые (хлорофиллы А и В), желто-оранжевые (ксантофилл и каротин) и др., ферменты, синтезирующие АТФ и переносчики электронов.
Рибосомы относят к немембранным органеллам клетки. На рибосомах осуществляется соединение аминокислотных остатков в полипептидные цепочки (синтез белка). Рибосомы очень малы и многочисленны. Каждая рибосома состоит из двух частей: малой и большой субъединиц. В первую входят молекулы белка и одна молекула рибосомальной РНК (р–РНК), во вторую - белки и три молекулы р–РНК (рис. 38). Белок и р–РНК по массе в равных количествах участвуют в образовании рибосом. Р–РНК синтезируется в ядрышке. В синтезе белка, кроме рибосом, принимают участие матричная РНК (м–РНК) и транспортная РНК (т–РНК). М–РНК несет генетическую информацию о синтезе белка от ядра. Эта информация закодирована в последовательном расположении нуклеотидов в молекуле м–РНК. М–РНК присоединяется к поверхности малой субъединицы. Т–РНК доставляет из цитоплазмы к рибосоме необходимые аминокислоты, из которых строится полипептидная цепь. В растущей полипептидной цепи каждая аминокислота занимает соответствующее место, что определяет качество синтезируемого белка. В процессе синтеза белка рибосома перемещается вдоль м–РНК.
В синтезе одной полипептидной цепочки участвуют много рибосом, соединенных последовательно друг с другом м–РНК. Такой комплекс из рибосом называют полирибосомой. Рибосомы удерживают в нужном положении аминокислоты, м–РНК, т–РНК до тех пор, пока между соседними аминокислотами не образуется пептидная связь.
Рибосомы могут свободно находиться в цитоплазме или быть связанными с эндоплазматической сетью, входя в состав шероховатой ЭПС.
Белки, образовавшиеся на рибосомах, соединенных с мембраной ЭПС, обычно поступают в цистерны ЭПС. Белки, синтезируемые на свободных рибосомах, остаются в гиалоплазме. Например, на свободных рибосомах синтезируется гемоглобин в эритроцитах. В митохондриях, пластидах и клетках прокариот также присутствуют рибосомы.
К немембранным органеллам относят микротрубочки и микрофиламенты. Микротрубочки – тончайшие трубочки диаметром 24 нм, стенки которых образованы белком тубулином. Глобулярные субъединицы этого белка располагаются по спирали. Микротрубочки определяют направление перемещения внутриклеточных компонентов, в том числе расхождение хромосом к полюсам клетки при делении ядра. Они участвуют в образовании «цитоскелета». Микрофиламенты – тонкие белковые нити диаметром 6 нм, состоят из белка актина, близкого тому, который содержится в мышцах. Эти нити, как и микротрубочки, являются элементами «цитоскелета». Они образуют кортикальный слой под плазматической мембраной. Кроме микротрубочек, присутствующих в цитоплазме, в клетке имеются микротрубочки, формирующие центриоли клеточного центра, базальные тельца, реснички и жгутики.
Клеточный центр располагается около ядра и состоит из парных центриолей и центросферы.
Центриоли характерны для животных клеток, их нет у высших растений, низших грибов и некоторых простейших. Центриоли окружены зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы (центросферы). Основу центриолей составляют девять триплетов микротрубочек (9+0), расположенных по окружности, и образующих полый цилиндр. Триплеты микротрубочек по кольцу объединены фибриллами. Радиальные фибриллы от каждого триплета отходят к центру, где они соединяются друг с другом.
В интерфазных клетках присутствуют две центриоли, расположенные под прямым углом друг к другу.
Перед делением ядра в синтетическом периоде центриоли удваиваются. В начале митоза к полюсам клетки направляются по две центриоли. Они принимают участие в формировании веретена деления, состоящего из микротрубочек. Центриоли участвуют в организации цитоплазматических микротрубочек.
В цитоплазме клеток присутствуют включения - непостоянные компоненты, выполняющие функцию запаса питательных веществ ( капли жира, глыбки гликогена), различных секретов, подготовленных к выведению из клетки. К включениям относят некоторые пигменты (гемоглобин, липофуцин) и другие. Включения синтезируются в клетке и используются в процессе обмена.
№7
Организация наследственного материала в клетке
В 1928 году Гриффит изучал безкапсульные неверулентные (т.е. не вызывающие заболевания) пневмококки и верулентные в полисахариде капсулы, вызывающие воспаление лёгкого. Опыты проводились для создания вакцины.
При инъекции мышам живых безкапсульных бактерий они выживали. А при введении живых капсульных – умирали.
При введении убитых при нагревании и живых безкапсульных пневмококков мыши погибали, а их них выделяли живых капсульных бактерий.
Таким образом способность образовывать капсулу перешла от одних бактерий к другим. Фактором, превращающим некапсульные пневмококки в патогенные, является ДНК, а само явление называется трансформацией.
Химическая природа ДНК была установлена Эвери, Макмедом, Маккарти в 1944 году.
Впервые ДНК из гноя выделил биохимическим методом швейцарский учёный Ф. Мишер в 1869 году.
ДНК – это полимер, мономером которого является нуклеотид. Нуклеотид состоит из азотистого основания, пуринового (аденин или гуанин, пуринового или перимединового), дизоксирибозы и остатка фосфорной кислоты.
Эдвин Чоргаф обнаружил, что количественное содержание гуанина равно содержанию цитозина, а содержание аденина равно содержанию тимина, и сформулировал правило Чаргафа:
отношение суммы Аденина и Гуанина равно сумме Тимина и Цитозина
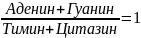
Основную структуру ДНК установили Уотсон и Крик:
ДНК – это двойная спираль, 10 пар оснований составляет полный оборот (3600). Основание находится внутри спирали, а их фосфатная группа находится снаружи. Таким образом первичная структура ДНК представляет собой цепочку нуклеотидов, в которой они связанны фосфодиэфирными связями.
Вторичная структура ДНК – представлена двойной цепочкой, в которой азотистые основания связанны водородными связями. Между Аденином и Тимином двойная связь, а между Гуанином и Цитозином – одинарная.
Третичная структура ДНК – это лево- или правозакрученная спираль.
Репликация.
Основное свойство ДНК – способность к репликации.
Значение репликации – обеспечение потомков полной и точной генетической информацией. Этот процесс был изучен в лаборатории А. Корнберга в 1955-57 году.
Репликация основывается на 3х принципах:
Комплиментарность – азотистые основания своей пространственной структурой должны дополнять друг друга.
Полуконсервативность – каждая молекула состоит из 1 цепи исходной родительской информации, а вторую синтезирует заново.
Антиполярность.
Для того, чтобы начался процесс репликации суперспирализованная ДНК должна быть релаксированна и цепи выпрямлены. Участок расхождения цепи ДНК называют репликативной вилкой. Ферментативный комплекс, осуществляющий репликацию может работать только в одном направлении – от 5’ к 3’ концу. Поэтому только одна из новых цепей будет ситезироваться непрерывно и называется лидирующей. Вторая цепь будет синтезироваться в виде фрагментов – фрагменты Оказаки, которые затем сшиваются в единую цепь. Синтез таких фрагментов идёт на фрагменте РНК (РНК-затравка или праймер), который в последствии удаляется.
Единицей репликации является репликон.
Ферментативный комплекс, участвующий в репликации ДНК:
ДНК-геликаза – расплетает двойную спираль.
Белки-дестабилизаторы – расплетают участки ДНК, делая её доступной для других ферментов.
ДНК-топоизомераза – разрывает фосфодиэфирные связи.
РНК-проймаза – синтезирует РНК-затравку.
ДНК-полимераза – основной элемент репликации. Синтезирует лидирующую цепь и фрагменты Оказаки.
ДНК-липаза – сшивает фрагменты Оказаки в единую цепь.
Репликация у прокариот и эукариот протекает не одинарно. У прокариот имеется одна кольцевая ДНК и она имеет только 1ну точку начала репликации, она же является точкой её окончания. Таким образом репликативная вилка проходит последовательно всю ДНК и функционирует как единственный репликон. Скорость полимеризации 500-1000 нуклеотидов.
ДНК эукариот имеет полирепликативное строение. Скорость полимеризации 50-100 нуклеотидов. Высокая точность обеспечивается специальными механизмами, которые осуществляют коррекцию.
Механизмы коррекции:
Механизм самокоррекции – осуществляется ДНК-полимеразой. Самокоррекция заключается в отщеплении ошибочно включённого в цепь ДНК нуклеотида, не спаренного с матрицей. Следствием самокоррекции снижается механизм ошибок в 10 раз.
Эксцизионная (дореплекативная) репарация – осуществляется специфическими ферментами. Искажение последовательностей нуклеотидов в одной из цепей обнаруживается ими, затем соответствующий участок удаляется и замещается новым, синтезированным на второй комплиментарной цепи ДНК.
Пострепликативная репарация – осуществляется путём рекомбинации (обмена фрагментами) между двумя вновь образованными двойными спиралями ДНК. Пример: возникновение тиминовых димеров, когда они не устраняются самопроизвольно под действием видимого света (световая репарация) или в ходе дорепликативной эксцизионной репарации. Ковалентные связи, возникающие между рядом стоящими остатками тимина делают их не способными к связыванию с комплиментарными нуклеотидами. В результате во вновь синтезируемой цепи возникают бреши (разрывы), узнаваемые ферментами репарации. Восстановление целостности новой полинуклеотидной цепи одной из дочерних ДНК осуществляется благодаря рекомбинации с соответствующей ей нормальной материнской цепью другой дочерней ДНК. Образовавшийся в материнской цепи пробел заполняется путём синтеза на комплиментарной ей полинуклеотидной цепи.
Если в наследственном материале слишком много повреждения и част из них не ликвидируется, включается система индуцируемых (побуждаемых) ферментов репарации (SOS-система). Эти ферменты заполняют бреши, восстанавливая целостность синтезируемых полинуклеотидных цепей без точного соблюдения принципа комплиментарности. Поэтому иногда сами процессы репарации могут служить источником стойких изменений в структуре ДНК (мутаций).
У прокариот большая часть генома представлена кодирующими последовательностями генотипа, которые несут геномную информацию.
У большинства высших эукариот в организации материала наблюдается избыточность ДНК. Их геном представлен:
Часто повторяющиеся последовательности – представлены в гаплоидном наборе 106 и более копиями. Эти последовательности не участвуют в синтезе РНК и называются сателитными.
Умеренно-повторяющиеся последовательности – представлены 102-105 копиями. В них входят гены, кодирующие структуру гистоновых белков тРНК и рРНК.
Уникальные последовательности – существуют в единственном числе или в 2-3х копиях. К ним относятся все гены, кодирующие структуры основных белков. С них копируются всё мРНК.
№8
Реализация биологической информации в клетке (транскрипция)
Всю последовательность процессов, происходящих при синтезе белковых молекул, можно объединить в 3 этапа:
Транскрипция
Процессинг
Трансляция
Структурными единицами наследственной информации являются гены участки молекулы ДНК, кодирующие синтез определенного белка.
Транскрипция (от лат. transcriptio – переписывание) – процесс синтеза молекулы и–РНК на молекуле ДНК, выступающей в роли матрицы. Молекула ДНК на участке гена раскручивается, и списывание информации происходит с одной из двух нитей молекулы ДНК, называемой кодогенной. Сборку молекулы и–РНК по принципу комплементарности осуществляет фермент – РНК–полимераза. Скорость сборки достигает 50 нуклеотидов в секунду. Списывание происходит только с части молекулы ДНК, называемой геном, и длина молекулы и–РНК в сотни раз короче, чем ДНК. Некоторые участки и–РНК не несут информацию о будущей молекуле белка. Их присутствие связано с особенностями строения генов и механизма транскрипции. Эти участки молекулы и–РНК, называемые интронами, необходимо удалить.
II. Процессинг – процесс созревания молекулы информационной РНК, сопровождающийся удалением интронов, участков, не несущих информацию о последовательности аминокислот в синтезируемом белке, и сращиванием (сплайсингом) остающихся фрагментов (экзонов, т.е. кодирующих последовательностей). Поэтому длина созревшей и направляющейся к рибосомам молекулы и–РНК оказывается короче первоначальной. Эту РНК называют матричной (м–РНК).
III. Трансляция (от лат. translatio – перевод) – синтез полипептидных цепей белков по матрице м–РНК на рибосомах.
Аминокислоты, из которых синтезируются белки, доставляются к рибосомам с помощью специальных транспортных РНК (т–РНК). Молекулы т–РНК, состоящие из 85–100 нуклеотидов, способны сворачиваться таким образом, что напоминают по форме лист клевера. В клетке присутствует около 40 молекул т–РНК. На вершине «листа» т–РНК имеется триплет, называемый антикодоном. Он комплементарен нуклеотидам кодона м–РНК. К основанию молекулы т–РНК присоединяется соответствующая аминокислота, та, которую кодирует триплет, комплементарный антикодону. Этот процесс осуществляется с помощью фермента – кодазы, с затратой энергии, получаемой при расщеплении молекулы АТФ. Трансляция состоит из трех последовательных фаз – инициации, элонгации и терминации.
Инициация. На этом этапе происходит сборка всего комплекса, участвующего в синтезе молекулы белка. Происходит при присоединенииферментативного комплекса РНК-полимеразы к промотору, при этом происходит расплетение двойной спирали ДНК.
2. Элонгация. В молекуле любой мРНК есть участок, комплементарный рРНК – малой субъединицы рибосомы и специфически ею управляемый. Рядом с ним находится инициирующий стартовый код он АУТ, кодирующий аминокислоту метионин. На рибосоме имеются два участка для связывания двух молекул тРНК. В одном участке, называемым пептидильным, уже находится первая тРНК. Это всегда одна и та же тРНК, несущая аминокислоту метионин (I). С него начинается синтез любой молекулы белка. Во второй участок рибосомы — аминоацильный поступает вторая молекула т–РНК и присоединяется к своему кодону (II). Между метионином и второй аминокислотой образуется пептидная связь. Вторая тРНК перемещается вместе со своим кодоном мРНК в пептидильный центр. Перемещение тРНК с полипептидной цепочкой из аминоацильного участка в пептидильный сопровождается продвижением рибосомы по м–РНК на шаг, соответствующий одному кодону. Этот этап требует затраты энергии. тРНК, доставившая метионин, возвращается в цитоплазму. Аминоацильный центр освобождается. В него поступает новая тРНК, связанная с аминокислотой, зашифрованной очередным кодоном (III). Между третьей и второй аминокислотами образуется пептидная связь, и третья тРНК вместе с кодоном м—РНК вновь перемещается в пептидильный центр. Таким образом, в растущей белковой молекуле аминокислоты оказываются соединенными в той последовательности, в которой расположены шифрующие их кодоны в мРНК. Процесс элонгации, удлинения белковой цепи, продолжается до тех пор, пока в рибосому не попадет один из трех кодонов, не кодирующих аминокислоты. Это триплеты терминации: УАА, УГА, УАГ. Ни одна из тРНК не может занять место в аминоацильном центре.
Терминация – прекращение роста цепи. Происходит на специальных участках – терминаторах, так как гены прокариот состоят из кодирующих последовательностей, несущих генетическую информацию ,первичные транскрипты мРНК могут сразу выполнять роль матрицы при трансляции. У эукариот транскрипция и трансляция разделены по времени и топографии.
Гены эукариот содержат некодирующие последовательности – интроны. Поэтому первичный транскрипт имеет большие размеры, чем необходимо для трансляции и называется ядерной РНК.
Она является точной копией транскрибированного участка ДНК. Эта молекула в дальнейшем подвергается процесингу (формированию зрелых мРНК), в ходе которой происходит сплайсинг (вырезание интронов и сшивание экзонов). В результате формируется мРНК, состоящая только из кодирующих последовательностей.
Ферментативный комплекс РНК-полимеразы, принимающий участие в транскрипции:
РНК-полимераза-1. Отвечает за транскрипцию генов рРНК.
РНК-полимераза-2. Отвечает за транскрипцию гетерогенной ядерной РНК.
РНК-полимераза-3. Отвечает за синтез рРНК и тРНК.
РНК-полимераза митохондрий (и пластид).
№9
Реализация биологической информации в клетке (трансляция)
Трансляция – это процесс воплощения генетической информации мРНК в структуру полипептида. Зрелая мРНК выходит яз ядра в цитоплазму, где соединяется с рибосомой.
Инициация трансляции начинается со стартового кодона (АУГ), который локализуется в смысловой части гена кодирующую аминокислоту метиламин. Аминокислоты доставляются к рибосоме тРНК, специальной для каждого вида аминокислот. Участок рибосомы, куда доставляется аминокислоты называется аминоацильный (α-участок).
Участок рибосомы, где происходит рост полипептидной цепи, называется пептидильным.
Этап элонгации связан с последовательным включением аминокислот в рост полипептидной цепи.
Этапы элонгации:
Связывание антикодона тРНК, несущей аминокислоту с кодоном мРНК в свободном α-участоке на рибосоме.
Образование пептидной связи между аминокислотами, когда в α- и ∏-участке находится тРНК с аминокислотами.
Транслокация. тРНК ∏-участка открывается от аминокислоты и покидает рибосому. тРНК из α-участка переходит в ∏-участок и рибосома перемещается вдоль мРНК на 1 кодон.
Терминация трансляции происходит, когда на мРНК в α-участке будет 1на из 3х стопкодонов (УАА, УАГ, АГА).
РНК
Рибонуклеиновые кислоты бывают нескольких видов. Есть рибосомальная, транспортная и информационная РНК. Нуклеотид РНК состоит из одного из азотистых оснований (аденина, гуанина, цитозина и урацила), углевода - рибозы и остатка фосфорной кислоты. Молекулы РНК - одноцепочковые.
Рибосомальная РНК (рРНК) в соединении с белком входит в состав рибосом. Р-РНК составляет 80% от всей РНК в клетке. На рибосомах идет синтез белка.
Информационная РНК (и-РНК) составляет от 1 до 10% от всей РНК в клетке. По строению и-РНК комплементарна участку молекулы ДНК, несущему информацию о синтезе определенного белка. Длина и-РНК зависит от длины участка ДНК, с которого считывали информацию. иРНК переносит информацию о синтезе белка из ядра в цитоплазму.
Транспортная РНК (тРНК) составляет около 10% всей РНК Она имеет короткую цепь нуклеотидов и находится в цитоплазме. тРНК присоединяет определенные аминокислоты и подвозит их к месту синтеза белка к рибосомам. ТРНК имеет форму трилистника. На одном конце находится триплет нуклеотидов (антикодон), кодирующий определенную аминокислоту. На другом конце имеется триплет нуклеотидов, к которому присоединяется аминокислота. При комплементарности триплета тРНК (антикодона) и триплета иРНК (кодона), аминокислота занимает определенное место в молекуле белка.
РНК находится в ядрышке, в цитоплазме, в рибосомах, в митохондриях и пластидах. В природе есть еще один вид РНК. Это вирусная РНК. У одних вирусов она выполняет функцию хранения и передачи наследственной информации. У других вирусов эту функцию выполняет вирусная ДНК.
№10
Клетка как открытая саморегулирующаяся биологическая система
Аденозинмонофосфорная кислота (АМФ) входит в состав всех РНК. При присоединении еще двух молекул фосфорной кислоты (Н3РО4) АМФ превращается в аденозинтрифосфорную кислоту (АТФ) и становится источником энергии, необходимой для биологических процессов, идущих в клетке.
Схема передачи энергии с помощью АТФ из реакций, в результате которых энергия освобождается (экзотермические реакции), в реакции, потребляющие эту энергию (эндотермические реакции). Последние реакции очень разнообразны: биосинтез, мышечные сокращения и т.д. Аденозинтрифосфорная кислота (АТФ) состоит из азотистого основания аденина, сахара — рибозы и трех остатков фосфорной кислоты. Молекула АТФ очень неустойчива и способна отщеплять одну или две молекулы фосфата с выделением большого количества энергии, расходуемой на обеспечение всех жизненных функций клетки (биосинтез, трансмембранный перенос, движение, образование электрического импульса и др.). Связи в молекуле АТФ называют макроэргическими. Отщепление концевого фосфата от молекулы АТФ сопровождается выделением 40 кДж энергии. Синтез АТФ происходит в митохондриях.
Энергетический обмен (диссимиляция)
представляет собой совокупность реакций расщепления органических веществ, сопровождающихся выделением энергии. В зависимости от среды обитания энергетический обмен может протекать в 2а или в 3и этапа.
У большинства организмов – аэробов, живущих в кислородной среде, в ходе диссимиляции осуществляется 3и этапа: подготовительный, бескислородный и кислородный, в процессе которых органические вещества расщепляются до неорганических.
Подготовительный этап. На этом этапе происходит ферментативное расщепление сложных органический соединений на более простые (белков – до аминокислот, жиров – на глицерин и жирные кислоты, полисахаридов – до моносахаридов, нуклеиновых кислот – до нуклеотидов). Распад органических субстратов пищи происходит на разных уровнях желудочно-кишечного тракта многоклеточных организмов. Внутриклеточное расщепление происходит под действием гидролитических ферментов лизосом. Высвобождающаяся при этом энергия рассеивается в виде теплоты, а образующиеся малые органические молекулы могут подвергаться дальнейшему расщеплению или использоваться клеткой как «строительный материал» для синтеза собственных органических соединений.
Неполное расщепление (бескислородный этап) – осуществляется непосредственно в цитоплазме клетки, в присутствии кислорода не нуждается и заключается в дальнейшем расщеплении органических субстратов. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное расщепление глюкозы называют гликолизом.
Гликолиз – многоступенчатый ферментативный процесс превращения шестиуглеродной глюкозы в 2е трёхуглеродные молекулы пировиноградной кислоты (пирувата, ПВК) С3Н4О3.
В ходе реакции гликолиза выделяется большое количество энергии – 200 кДж/моль. Часть этой энергии (60%) рассеивается в виде теплоты, а остальное (40%) используется на синтез АТФ. В результате гликолиза одной молекулы глюкозы образуется по 2е молекулы ПВК, АТФ и воды, а так же атомы водорода, которые запасаются клетной в форме НАД*Н, то есть в составе переносчика – никонинамидадениндинуклеотида. Суммарная формула гликолиза имеет следующий вид:
С6Н12О6+2АДФ+2Ф+2НАД+ → 2С3Н4О3+2АТФ+2Н2О+2НАД*Н
дальнейшая судьба продуктов гликолиза – пирувата и водорода в форме НАД*Н – может складываться по-разному. У дрожжей или в клетках растений при недостатке кислорода происходит спиртовое брожение – ПВК восстанавливается до этилового спирта:
(пируват) СН3СОСООН → СО2+СН3СОН (укс. альдегид)
(укс. альдегид) СН3СОН+НАД*Н → С2Н5ОН (этил. спирт)+НАД+
В клетках животных, испытывающих временный недостаток кислорода, например в мышечных клетках человека при чрезмерной физической нагрузке, а так же у некоторых бактерий происходит молочнокислое брожение, при котором пируват восстанавливается до молочной кислоты:
(пируват) СН3СОСООН+НАД*Н → С3Н6О3 (лактат)+НАД+
Полное окисление (дыхание или кислородный этап), протекает при обязательном участии кислорода. Аэробное дыхание представляет собой цепь реакций, контролируемых ферментами внутренней мембраной и матриксом митохондрий. Попав в митохондрию, ПВК взаимодействует с ферментами матрикса и образует: диоксид углерода, который выводится из клетки; атомы водорода, которые в составе переносчиков направляются к внутренней мембране; ацетилкофермент А (ацетил-КоА), который вовлекается в цикл трикарбоновых кислот (цикл Кребса). Цикл Кребса – это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуется 2е молекулы СО2 , молекула АТФ и 4ре пары водорода, передаваемые на молекулы переносчики – НАД и ФАД. Суммарную реакцию гликолиза и цикла Кребса можно определить в следующем виде:
С6Н12О6+6Н2О → 6СО2+4АТФ+12(НАД*Н2+ФАД*Н2)
Итак, в результате бескислородного этапа диссимиляции и цикла Кребса молекула глюкозы расщепляется до неорганического диоксида углерода, а высвободившаяся при этом энергия частично расходуется на синтез АТФ, но в основном сберегается в нагруженных электронами переносчиках НАД*Н2 и ФАД*Н2 . Белки переносчики транспортируют атомы водорода к внутренней мембране митохондрий, где передают их по цепи встроенных в мембрану белков. Транспорт частиц по цепи переноса осуществляется таким образом, что протоны остаются на внешней стороне мембраны и накапливаются в межмембранном пространстве, превращая его в Н+ резервуар, а электроны передаются на внутреннюю поверхность внутренней митохондриальной мембраны, где соединяются в конечном итоге с кислородом:
О2+е- → О2-
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно, а снаружи – положительно, так что между её поверхностями создаётся разность потенциалов. Когда разность потенциалов достигает критического уровня, положительно заряженные частицы Н+ силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду:
 О2+2Н+
→ Н2О
О2+2Н+
→ Н2О
При этом энергия транспортирующихся ионов водорода используется для фосфорилирования АДФ в АТФ:
АДФ+Ф → АТФ
Известно, что 55% энергии запасается в химических связях АТФ, а 45% - рассеивается в виде теплоты. Синтез АТФ в процессе клеточного дыхания тесно сопряжён с транспортом ионов по цепи переноса и называется окислительным фосфорилированием.
2С3Н6О3+36АДФ+36Ф+6О2 →(+НАД)→ 6СО2+36АТФ+6Н2О
№11
Существование клеток во времени (клеточный цикл)
Увеличение числа клеток происходит путем деления исходной клетки. Обычно делению клеток предшествует редупликация хромосомного аппарата, синтез ДНК. Время существования клетки от деления до следующего деления или смерти называют клеточным (жизненным) циклом. В течение жизни клетки растут, дифференцируются, выполняют определенные функции, размножаются, гибнут. В клеточном цикле можно выделить митотический цикл, включающий подготовку клеток к делению и само деление. В жизненном цикле есть периоды, когда клетки выполняют определенные функции.
В, организме высших позвоночных не все клетки постоянно делятся. Есть специализированные клетки, потерявшие способность к делению (нейтрофилы, базофилы, эозинофилы, нервные клетки). Другие клетки способны постоянно делиться. Они обнаружены в обновляющихся тканях (эпителиальных), в кроветворных органах. Например, клетки покровного эпителия, кроветворные клетки костного мозга могут постоянно делиться, заменяя погибшие. Многие клетки, не размножающиеся в обычных условиях, начинают делиться в процессе восстановления после повреждения органа и репаративной регенерации органов и тканей. Клетки, находящиеся в клеточном цикле, содержат различное количество ДНК, в зависимости от стадии этого цикла. Мужские и женские половые клетки имеют гаплоидный набор хромосом (n) и количество ДНК (с). При оплодотворении происходит слияние этих клеток, в результате чего образуется диплоидная клетка с 2n набором хромосом и 4с количеством ДНК. Удвоение ДНК происходит в синтетическом периоде интерфазы. Клетки к делению приступают только после этого периода.
Подготовка клетки к делению
В клеточном цикле можно выделить собственно митоз и интерфазу, включающую пресинтетический (постмитотический) — G1 период, синтетический (S) период и постсинтетический (премитотический) - G2 период.
Подготовка клетки к делению происходит в интерфазе. Пресинтетический период интерфазы - самый длительный. Он может продолжаться у эукариот от 10 часов до нескольких суток.
В пресинтетическом периоде (G1), наступающем сразу после деления, клетки имеют диплоидный (2n) набор хромосом и 2с генетического материала ДНК. В этот период начинается рост клеток, синтез белков, РНК. Происходит подготовка клеток к синтезу ДНК (S-период). Повышается активность ферментов, участвующих в энергетическом обмене.
В S-периоде (синтетическом) происходит репликация молекул ДНК, синтез белков - гистонов, с которыми связана каждая нить ДНК. Синтез РНК увеличивается соответственно количеству ДНК. При репликации две спирали молекулы ДНК раскручиваются, рвутся водородные связи, и каждая становится матрицей для воспроизводства новых цепей ДНК. Синтез новых молекул ДНК осуществляется при участии ферментов. Каждая из двух дочерних молекул обязательно включает одну старую и одну новую спираль. Новые молекулы идентичны старым. Такой способ репликации называют полуконсервативным. В S-периоде начинается удвоение центриолей. Каждая хромосома состоит из двух сестринских хроматид, содержит ДНК 4с. Число хромосом не меняется (2n). Продолжительность синтеза ДНК - S-период митотического цикла - длится 6-12 часов у млекопитающих.
В постсинтетический период (G2) происходит синтез РНК, накапливается энергия АТФ, необходимая для деления клетки, завершается удвоение центриолей, митохондрий, пластид, синтезируются белки, из которых строится ахроматиновое веретено деления, заканчивается рост клетки. Ни содержание ДНК (4с), ни число хромосом (2n) не изменяется.
Продолжительность этого периода – 3 - 6 часов. Длительность клеточного цикла разная у разных клеток, но постоянна для данной ткани. Например, в культуре раковых клеток человека длительность G1-периода равна 8,5 часов, S - 6,2 часа, G2 - 4,6 часов. Длительность митоза составляет 0,6 часа. Весь клеточный цикл длится 19,9 часов.
№12
Механизмы клеточной пролиферации (митоз)
Существуют три способа деления клетки: митоз, амитоз, мейоз. Митоз - mitos (греч. - нити) - непрямое деление клетки.
Митоз состоит из четырех фаз: профазы, метафазы, анафазы, телофазы.
Профаза занимает — 0,60 времени от всего митоза, метафаза — 0,05 времени, анафаза — 0,05 и телофаза — 0,3 времени всего митоза. Длительность митоза различна у разных клеток, но не менее 10 минут. В интерфазном ядре хромосомы под световым микроскопом не видны.
В профазе увеличивается объем ядра. Хромосомы спирализуются, становятся видимыми, укорачиваются, утолщаются. Видно, что они состоят из двух хроматид, соединенных центромерой. Центриоли расходятся к полюсам клетки. Формируется веретено деления. К концу профазы ядрышки и ядерная оболочка растворяются, и хромосомы оказываются в цитоплазме. Профаза - самая продолжительная фаза митоза. В профазе набор хромосом равен 2n, и количество ДНК равно 4с.
В метафазе спирализация достигает максимума, хромосомы располагаются в экваториальной плоскости веретена, образуя метафазную пластинку. Сестринские центромеры и хроматиды обращены к противоположным полюсам. Митотическое веретено полностью сформировано и состоит из нитей, соединяющих полюса с центромерами хромосом. Отчетливо видно, что хромосомы состоят из двух хроматид, соединенных в области центромеры. Четко видны число и форма хромосом, что позволяет сосчитать их и изучить строение. Метафаза очень короткая.
В анафазе центромеры разъединяются, хроматиды (дочерние хромосомы) становятся самостоятельными. Нити веретена деления, прикрепленные к центромерам, тянут дочерние хромосомы к полюсам клетки. Движение хромосом обеспечивается взаимодействием центромерных участков хромосом с микротрубочками веретена деления. В клетке находятся два диплоидных набора хромосом. Анафаза очень короткая.
Митоз заканчивается телофазой. Хромосомы, состоящие из одной хроматиды, находятся у полюсов клетки. Они деспирализуются и становятся невидимы. Образуется ядерная оболочка, нити ахроматинового веретена распадаются. В ядре формируется ядрышко. Происходит деление цитоплазмы (цитотомия и цитокинез) и образование двух дочерних клеток. В клетках животных цитоплазма делится путем перетяжки, впячиванием цитоплазматической мембраны от краев к центру. В клетках растений - в центре образуется мембранная перегородка, которая растет по направлению к стенкам клетки. После образования поперечной цитоплазматической мембраны у растений образуется целлюлярная стенка.
В результате митоза происходит точное распределение генетического материала между двумя дочерними клетками. Обе дочерние клетки получают диплоидный набор хромосом. Митоз обеспечивает поддержание постоянства числа хромосом в ряду поколений и служит клеточным механизмом процессов роста, развития организма, регенерации, бесполого размножения. При нарушении хода митоза, происходящего под действием некоторых ядов, наблюдается нерасхождение хромосом, нарушение их строения, повреждение веретена деления. Вследствие повреждений имеют место различные мутации.
Амитоз
Амитоз - прямое деление клетки, при котором ядро находится в интерфазном состоянии. Хромосомы не выявляются. Веретено деления не образуется. Амитоз приводит к появлению двух клеток, но очень часто в результате амитоза возникают двуядерные и многоядерные клетки. Амитотическое деление начинается с изменения формы и числа ядрышек. Крупные ядрышки делятся перетяжкой. Вслед за делением ядрышек происходит деление ядра. Ядро может делиться перетяжкой, образуя два ядра или имеет место множественное разделение ядра, его фрагментация. Ядра могут быть неравной величины. Амитоз встречается в отживающих, дегенерирующих клетках, неспособных дать новые жизнеспособные клетки. В норме амитотическое деление ядер встречается в зародышевых оболочках животных, в фолликулярных клетках яичника. Амитотически делящиеся клетки встречаются при различных патологических процессах (воспаление, злокачественный рост и др.).
№13
Хромосомный уровень организации наследственного материала
Хромосома – это уровень организации наследственного материала в виде нуклеотидного комплекса. Первое упоминание было в 1880 г., когда Флеминг при обследовании роговицы глаза человека обнаружил хроматиновые тела.
Термин хромосова введён в 1888 г.
Во фракцию хроматина кроме ДНК входит
Белки (60 – 70% сухой массы), которые в свою очередь делятся на:
Гистоновые (это сильноосновные белки, щёлочность которых обуславливается основными аминокислотами (лизином и аргинином).
Препарат гистонов состоит из 5 фракций:
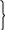 - Н1
- Н1
-Н2А - умеренно богаты лизином
-Н2В
 -Н3
- умеренно богаты
аргинином
-Н3
- умеренно богаты
аргинином
-Н4
Все гистоны, кроме Н1 сходны по своей структуре и содержатся во всех клетках человека примерно в равных количествах, а Н1 в 2 раза меньше.
Гистоны синтезируются на полисомах цитоплазмы, мигрируют в ядро, связываются с ДНК и остаются с ней на протяжении ряжа клеточных делений.
Негистоновые белки (составляют 20% от массы гистоновых)
Во фракцию негистоновых белков входят:
- ферменты репликации
- ферменты репарации
- транскрипции
- модификации ДНК
- белки регуляторы и т.д.
РНК (составляет от 0,2 – до 15% от содержания ДНК). Представлена всеми известными видами РНК, находящихся в процессе синтеза или созревания.
Лепиды и полисахариды (до 0,5%).
Интерфазная и митотическая хромосомы – это 2 варианта одной химической организации. Согласно исследованиям хромосома представлена нитями с различной степенью компактизации.
Уровни компактизации:
Нуклеосома – представляет собой структуру «Бусы на нитке», где фрагмент ДНК из 146 пар нуклеотидов делает виток вокруг поверхности гистоновой сердцевины (коры). Она представлена октаэдром гистонов Н2А, Н2В, Н3 и Н4. Диаметр нуклеосомы равен 10 нм. ДНК свободная от гистоновой называется линкерной или связующей (содержит 50 пар нуклеотидов).
Следовательно, 1 нуклеосомная единица = 146 + 50 = 196 пар нуклеотидов.
Гистон Н1 в формировании нуклеосомной субъединицы не участвует, а выполняет роль скрепки, т.е. располагаясь на ДНК соединяет 1 нуклеосому с другой.
Хромосомная фибрилла – несколько нуклеосом сближены в линейном порядке (модель соленоида). Диаметр равен 30 нм.
Хромонемный – хромосомные фибриллы объединятся с помощью негистоновых белков. Это уровень интерфазного хроматина.
Метафазная хромосома – достигается максимальная степень компактизации (диаметр равен 300 нм.).
Морфологическая неодородность хромосомы определяется изменчивостью её конденсации и спирализации. Проходя к началу митоза полную кондерсацию одни районы хроматина деспирализуются (эухроматин), а другие остаются в спирализованном состоянии (гетерохроматин).
Гетерохроматин не содержит структурных генов и является генетически инертным. Весь комплекс структурных генов находится в эухроматине.
Кариотип – это совокупность числа и величины морфологических хромосом, по которому можно идентифицировать данный набор хромосом.
Виды хромосом:
Метацентрическая – равноплечая.
Субметацентрическая – одно плечо немного короче другого.
Акроцентрическая – с коротким или едва заметным 2 плечом.
Телоцентрическая – место первичной перетяжки трудно различимо (в кариотипе человека их нет).
Для идентификации хромосом применяют способ окрашивания.
2 способа окрашивания:
Простое (рутинное) окрашивание - 2% ацетореином или ацетокормином. Окрашивает хромосому целиком равномерно и интенсивно по всей длине.
Дифференциальное окрашивание – при этом выявляется в хромосомах чередование тёмных и светлых сегментов, которые оказываются постоянными.
Это сделало возможным создание в 1971 году в Париже идеограммы хромосом человека. До это в 60х годах Денвере (США) была создана классификация, где хромосомы располагались и нумеровались согласно их длинны. В Париже добавилось их линейное описание.
Пары нумеруют с 1ой по 23ю. Патау предложил разбить их на 8 групп.
Группы:
A (с 1ой по 3ю пару) – 1я и 3я пары большие метацентрические, а 2я большая субметацентическая.
B (с 4ой по 5ю пару) – большие субметацентрическте.
C (с 6ой по 12ю пару) – средние субметацентрические. При рутинной окраске половые хромосомы неотличимы от хромосом этой группы.
D (с 13ой по 15ю пару) – средние акроцентрики. Все эти хромосомы имеют спутники.
Е (с 16ой по 18ю пару) – небольшие субметоцентрические хромосомы.
F (с 19ой по 20ю пару) – малые метацентрики.
(23я пара) – половые хромосомы. Мужская Y-хромосома является средней акроцентрической. У мужчин 11 акроцентрических хромосом.
№14
Размножение организмов: размножение как фундаментальное свойство живого
Размножение – способность живых организмов воспроизводить потомство. Обычно выделяют два основных типа размножения – бесполое и половое.
Бесполое размножение.
Осуществляется при участии лишь одной родительской особи. Особи дочернего поколения возникают из оной или группы клеток материнского организма. Наиболее широко бесполое размножение встречается среди прокариот, грибов и растений, но встречается и у различных видов животных. Основными формами бесполого размножения являются деление, спорообразование, фрагментация и вегетативное размножение.
У одноклеточных организмов в благоприятных условиях размножение осуществляется путём простого или множественного деления. При этом ядро один или несколько раз делится митозом с образованием двух или большего числа дочерних ядер, каждое из которых окружается цитоплазмой и формирует самостоятельных организм.
У многих растений (водоросли, мхи, папоротники), грибов и некоторых одноклеточных организмов на определённой стадии жизненного цикла образуются споры – мелкие гаплоидные клетки, покрытые плотной оболочкой и устойчивые к действию неблагоприятных факторов внешней среды. Большинство спор неподвижны и расселяются во внешней среде пассивно. Некоторые водоросли и грибы образуют подвижные зооспоры. При возникновении благоприятных условий среды споры прорастают , давая начало новому организму. Сам процесс называется спорообразованием.
В случаях, когда формирование нового организма осуществляется из группы клеток материнского организма, говорят о вегетативном размножении. Особенно широко встречается среди растений, у которых оно происходит за счёт частей вегетативных органов или специально предназначенных для этого структур – луковиц, корневищ, клубней и тд.
Различают так же почкование и фрагментацию. У некоторых грибов и животных от родительской особи отделяется небольшой участок тела – почка, из которой впоследствии развивается новый организм.
Фрагментацией называют разделение особи на две или несколько частей, каждая из которых растёт и достраивает новый организм. Основу фрагментации составляет способность организмов к регенерации.
Поскольку основным клеточным механизмом бесполого размножения является митоз, особи дочернего поколения являются точными копиями родительского организма.
Половое размножение
Половое размножение характеризуется наличием полового процесса, при котором происходит слияние гаплоидных половых клеток (гамет), образовавшихся в результате мейоза. Следствием этого является неповторимость каждой особи в любой популяции, размножающейся половым путем. Некоторые одноклеточные и многоклеточные организмы могут размножаться бесполым путем. Другие организмы, чтобы не погибнуть, периодически осуществляют половой процесс.
Разнообразны формы полового процесса у одноклеточных. Он может осуществляться по типу конъюгации, при которой не образуются специальные половые клетки, и по типу копуляции, когда особи приобретают половые различия, т.е. превращаются в гаметы и полностью сливаются, образуя зиготу. Конъюгация происходит у инфузорий при неблагоприятных условиях. Инфузории имеют два ядра: макронуклеус и микронуклеус. Макронуклеус отвечает за обменные процессы, микронуклеус принимает участие в половом процессе. При конъюгации две инфузории сближаются, между ними образуется цитоплазматический мостик. Макронуклеус растворяется, микронуклеус делится мейозом. В результате образуются четыре гаплоидных ядра, три из которых растворяются. Оставшееся ядро делится митозом. Образуются гаплоидные стационарное и мигрирующее ядра. Происходит обмен мигрирующими ядрами. После обмена в каждой из инфузорий мигрирующее и стационарное ядра сливаются, образуя синкарион (греч. syn - вместе, karyon - ядро), содержащий диплоидный набор хромосом. После конъюгации инфузории расходятся. Из синкариона формируются макро- и микронуклеусы. В результате конъюгации произошел обмен наследственной информацией, вследствие чего возникли новые комбинации генов, повышающие жизнеспособность особей.
Копуляция - половой процесс у одноклеточных организмов, при котором полностью сливаются копулирующие особи, выполняющие функции половых клеток (гамет). Копуляция может быть изогамной (греч. isos - равный, gsmos - брак), если особи, участвующие в копуляции, имеют одинаковые малые размеры, обе подвижны. Так размножается представитель колониальных жгутиковых – политома.
В анизогамной копуляции (греч. anisos - неравный, gamos -брак) участвуют две особи, одна из которых крупная и подвижная, а вторая мелкая и подвижная. Например, анизогамная копуляция характерна для колониальных жгутиковых пандорины. У пандорины могут сливаться при анизогамной копуляции большая и малая гаметы, или малая с малой, как при изогамной копуляции.
У колонии вольвокс из класса Жгутиковых происходит оогамная копуляция, при которой большая гамета неподвижна, а малая подвижна. У многих животных при половом размножении имеет место оогамная копуляция.
Основные этапы эволюции полового размножения
Эволюция размножения шла, как правило, в направлении от бесполых форм к половым, от изогамии к анизогамии, (Гаметы) от участия всех клеток в размножении к разделению клеток на соматические и половые, от наружного оплодотворения к внутреннему с внутриутробным развитием (Половая система) и заботой о потомстве. Темп размножения, численность потомства, частота смены поколений наряду с другими факторами определяют скорость приспособления вида к условиям среды. Например, высокие темпы размножения и частая смена поколений позволяют насекомым в короткий срок вырабатывать устойчивость к ядохимикатам. В эволюции позвоночных - от рыб до теплокровных - наблюдается тенденция к уменьшению численности потомства и увеличению его выживаемости.
№15
Гаметогенез
Гаметогенез – это процесс образования половых клеток называется гаметогенезом. Сперматозоиды образуются в результате сперматогенеза, яйцеклетки - овогенеза. Клетки зачаткового эпителия половых желез делятся последовательно митозом и мейозом. В результате этих делений образуются мужские гаметы (сперматозоиды) и женские гаметы (яйцеклетки). Они имеют гаплоидный набор хромосом и содержат наследственную информацию, необходимую для развития организма.
Сперматогенез.
Сперматогенез - это процесс образования мужских половых клеток. Происходит в извитых семенных канальцах, стенка которых образованна сперматогенным эпителием. Там находятся клетки на разных стадиях сперматогенеза и сопровождающие их клетки Сертоли.
Функция клеток Сертоли:
Трофическая (питание)
Гематотестикулярного барьера – пропускают дифференцированные клетки ближе к просвету канальца, препятствуют проникновению токсических веществ из крови к сперматогенному эпителию.
Участвуют в сперматогенезе – они синтезируют андроген-связывающий белок, которых переносит мужские половые гормоны к сперматогенным клеткам.
У половозрелых мужчин сперматогоний около 1 млд.
Различают:
А–сперматогонии - делятся митозом и остаются для поддержания популяции.
Б–перматогонии - делятся митозом, затеем дифференцируются в сперматоциты 1ого порядка (2n4c) и вступают в мейоз. После мейоза I образуется 2а сперматозоида второго порядка (n2c).
После мейоза II – 4ре сперматиды (nc). Они вступают в стадию формирования зрелого сперматозоида, при этом уменьшается количество цитоплазмы (она поглощается клетками Сертоли), образуется акросома с комплексом протеолитических ферментов. Появляется жгутик. Сохраняет жизнедеятельность в течении 1-2 суток. В 1 кубическом мм. семенной жидкости содержится около 100 млн. сперматозоидов.
Овогенез.
Овогенез - это процесс образования женских половых клеток. Происходит в корковом слое яичников. Стадия размножения женских половых клеток протекает во внутриутробном периоде с 3 по 7 месяц. При этом образуется примордикальная фолликула, каждая из которых покрыта одним слоем эпителия. Затем овогоний, прекративший размножение становится овоцитом (2n2c). После рождения большинство овоцитов дегенерируют, остальные вступают в мейоз I и после вступают в стадию пахитены (когда хромосомы принимают вид ламповых щёток). На ней мейоз останавливается на многие годы.
В период полового созревания под действие гормона гипофиза овоциты заканчивают мейоз I. При этом образуется овоцит II (n2c) и одно редукционное тельце. После этого созревающая яйцеклетка разрывает фолликул и выходит в брюшную полость, а от туда в маточную трубу.
Мейоз II завершается в маточной трубе при одном условии – если произойдёт оплодотворение.
В яичниках новорожденной девочки содержится около 2 млн. овоцитов, к периоду полового созревания их остаётся около 300 тыс., за весь репродуктивный период – примерно 400 яйцеклеток.
№16
Мейоз
Мейоз (от греч. meiosis - уменьшение) - процесс деления клеточного ядра с образованием четырех дочерних ядер, каждое из которых содержит вдвое меньше хромосом, чем исходное ядро. Мейоз состоит из двух слудующих друг за другом делений, сопровождающимися только одним удвоение ДНК. В мейозе I и II выделяют те же стадии, что и в митозу. Особый интерес представляет профаза I мейоза. В зависимости от перестройки её делят на 5 стадий:
Лептотена – стадия тонких нитей (хромосомы удвоены, но сестринские хроматиды трудноразличимы).
Зиготена – стадия сливающихся нитей. Это стадия прохождения конъюгации гомологичных хромосом (синапсис). При этом 2е хромосомы сближаются, образуя комплекс бивалент, состоящий из 4х хроматид. Число бивалент равно числу хромосом в гаплоидном наборе. На этой стадии образуется небольшое количество L-ДНК, состоящей из уникальных последовательностей нуклеотидов. Её специализированные участки имеют способность узнавать друг друга и закрепляют биваленты по типу застёжки молнии.
Пахитена – стадия толстых нитей. На это стадии происходит одно из главных событии мейоза – кроссинговер (обмен гомологичными участками между хромосомами). Результатом является рекомбинация скреплённых генов и образование отличных от исходных.
На этой стадии во время овогенеза активируется транскрипционная способность хромосом и они принимают вид ламповых щёток.
Диплотена – стадия двойных нитей. Происходит отталкивание гомологов друг от друга и они остаются связанными только в местах кроссинговера (хиазмах).
Диакинез – стадия обособления.
В метафазе I биваленты выстраиваются вдоль экватора клетки.
В анафазе I происходит расхождение гомологичных хромосом к противоположным полюсам клетки, причём расхождение гомологов каждой пары есть явление случайное.
Телофаза I. В телофазе первого мейотического деления число хромосом в каждой клетке становится гаплоидным. Хромосомы состоят из двух хроматид. Вследствие кроссинговера при образовании хиазм, хроматиды генетически не однородны. На короткое время образуется ядерная оболочка, хромосомы деспирализуются, ядро становится интерфазным. Затем у животной клетки начинается деление цитоплазмы, а у растительной клетки формирование клеточной стенки. У многих растений нет телофазы I, клеточная стенка не образуется, нет интерфазы II, клетки сразу переходят из анафазы I в профазу II.
Интерфаза II. Эта стадия есть только у животных клеток. Во время интерфазы между первым и вторым делением в S период не происходит редупликация молекул ДНК. Второе мейотическое деление называют эквационным. Оно похоже на митоз. Из хромосом, имеющих две хроматиды, образуются хромосомы, состоящие из одной хроматиды.
Профаза II. В профазе второго мейотического деления хромосомы утолщаются и укорачиваются. Ядрышко и ядерная оболочка разрушаются. Образуется веретено деления.
Метафаза II. В метафазе второго мейотического деления хромосомы выстраиваются вдоль экватора. Нити ахроматинового веретена отходят к полюсам. Образуется метафазная пластинка. Анафаза II. В анафазе второго мейотического деления центромеры делятся и тянут за собой к противоположным полюсам отделившиеся друг от друга хроматиды, называемые хромосомами. Телофаза II. В телофазе второго мейотического деления хромосомы деспирализуются, становятся невидимыми. Нити веретена исчезают. Вокруг ядер формируется ядерная оболочка. Ядра содержат гаплоидный набор хромосом. Происходит деление цитоплазмы и образование клеточной стенки у растений. Из одной исходной клетки образуются четыре гаплоидных клетки.
Значение мейоза
Поддержание постоянства числа хромосом. Если бы не возникало редукции числа хромосом при гаметогенезе, и половые клетки имели гаплоидный набор хромосом, то из поколения в поколение возрастало бы их число.
При мейозе образуется большое число новых комбинаций негомологичных хромосом.
В процессе кроссинговера имеют место рекомбинации генетического материала.
Практически все хромосомы, попадающие в гаметы, содержат участки, происходящие как первоначально от отцовской, так и от материнской хромосомы. Этим достигается большая степень перекомбинации наследственного материала. В этом одна из причин изменчивости организмов, дающая материал для отбора.
№17
Биологический аспект репродукции человека (оплодотворение)
Половой диморфизм – это различия меж особей разного пола, который проявляется на молекулярном и генетическом уровнях, по кариотипу и главному половому гормону. На организменном уровне имеются различия в показателях длинны волос трубчатых волос, в степени и характере оволосенения, строении гортани, в развитии мускулатуры, молочных желёз, распределении жировой ткани, строении половых органов.
Для участия в половом размножении в родительских организмах вырабатываются гаметы – клетки, специализированные к обеспечению генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы – клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуально развития.
У некоторых организмов зигота образуется в результате объединения гамет, не отличимых по строению. В таких случаях говорят о изогамии. У большинства видов по структурным и функциональным признакам половые клетки делятся на материнские (яйцеклетки) и отцовские (сперматозоиды). Как правило, яйцеклетки и сперматозоиды вырабатываются разными организмами.
Образование гамет обоих видов в одном организме называют гермафродитизмом (характерен для паразитов человека – плоских червей). Истинный гермафродитизм описан у человека. Чаще он развивается в результате нарушения эмбриогенеза при одинаковом наборе половых хромосом.
Оплодотворение
Оплодотворение происходит в несколько фаз.
Фаза I – сближение. Оно обеспечивается совокупностью, повышающая вероятность сближения гамет (скоординированность к наступлению оплодотворении. у обоих полов, поведение полов, избыточная продукция сперматозоидов, крупные яйцеклетки). Способствуют сближению и взаимодействию половых клеток: гиногамоны – выделяемые яйцеклеткой вещества, активизирующие движение сперматозоидов, вызывают их склеивание при встрече с андрогенами (реакция аглютинизации). Её биологический смысл – предохранение яйцеклетки от избытка сперматозоидов.
Фаза II – в ней происходит акросомная реакция. Наружная мембрана сперматозоида разрывается, высвобождаются протеолитические ферменты и растворяют оболочку яйцеклетки. Плазматические мембраны сливаются, цитоплазмы соединяются. В цитоплазму яйцеклетки переходят ядро и центриоль сперматозоида. Хвостовая часть рассасывается. Затем яйцеклетка активируется, меняется её потенциал и отслаивается её желточная оболочка и образуется оболочка оплодотворения (кортикальная реакция). Активация заканчивается началом синтеза белка.
Фаза III – сингамия. В ней выделяют:
Стадия 2х пронуклеусов – мужское ядро набухает, принимает вид профазного, за это время удваивается ДНК и мужской пронуклеус получает гаплоидный набор редуплецированных хромосом (n2c). Яйцеклетка в момент встречи со сперматозоидом находится в стадии мейоза, заблокированной с помощью специального фактора. После встречи со сперматозоидом, яйцеклетка активируется и блок снимается. Ядро яйцеклетки, закончившее мейоз, превращается в женский пронуклеус, так же приобретая набор хромосом n2c.
Стадия синкариона – слияние ядерного материала и образование зиготы.
Первое митотическое деление зиготы приводит к образованию двух клеток зародыша (бластомеров) с набором хромосом 2n2c в каждом.
Мужское и женское бесплодие
Женское бесплодие - неспособность зрелого женского организма воспроизвести потомство. Различают первичное и вторичное женское бесплодие. Первичным страдают женщины, которые никогда ранее не были беременны и которые при регулярной половой жизни с мужчиной без использования контрацептивов не могут забеременеть в течение года. Вторичное - это неспособность женщин, которые ранее были беременны, забеременеть снова.
Причины бесплодия
Основными причинами бесплодия являются:
воспалительные заболевания (в том числе половые инфекции)
непроходимость маточных труб
гормональные изменения
иммунологические факторы
Основной причиной женского бесплодия являются воспалительные заболевания половых органов. Часто заболевания, передающиеся половым путем , протекают безсимптомно и могут перейти в хроническую фазу, если лечение не было проведено вовремя. К бесплодию могут привести такие заболевания как хламидиоз и гонорея. Частым осложнением у женщин, которые перенесли сальпингит, эндометрит, гонорею или хламидиоз, может быть непроходимость маточных труб. Спайки не дают яйцеклетке попасть в матку и оплодотвориться. Гормональные изменения тоже могут быть причиной бесплодия. В идеале гормональная система отвечает за нормальный процесс овуляции. Если происходят какие-либо изменения в гормональной системе, то процесс овуляции будет проходить с нарушениями или вообще прекратится. При другом гормональном нарушении - снижении количества "прогестерона" - яйцеклетка будет оплодотворена, но не сможет закрепиться в матке. Самым неизученным фактором является иммунологический. В данном случае можно говорить о несовместимости, когда в женском организме вырабатываются "антитела" на сперму партнера, и оплодотворение становится невозможным.
Мужское бесплодие - это неспособность мужчины к производству потомства. Различают первичное бесплодие и вторичное. При первичном - когда от данного мужчины женщина не может забеременеть в течении года регулярной половой жизни. Это чаще связано с врожденной патологией (гипоспадия, эписпадия, вирусные заболевания и т.д.). При вторичном бесплодии мужчина ранее имел детей, однако приобретенные инфекции, хронический простатит и др. причины приводят к инфертильности. Наиболее частыми причинами бесплодия у мужчин являются: - воспалительные заболевания (половые инфекции)
травмы мошонки, операции на мошонке
эпидемический паротит (свинка)
длительное воздействие тепла (в том числе на мошонку)
гормональные изменения
иммунологические факторы
варикоцеле
Заболевания, передающиеся половым путем являются основной причиной мужского бесплодия. Особенно грозным заболеванием является гонорея. При этом заболевании просвет мочеиспускательного канала, а также семявыбрасывающего протока значительно сужается. Это приводит к бесплодию. Недостаток мужских гормонов (например тестостерона) а также избыток женских половых гормонов приводит к снижению количества сперматозоидов и снижению их подвижности. Температура яичек должна быть на 4 градуса ниже температуры тела. Слишком высокая температура нарушает образование сперматозоидов. Например, одно-единственное погружение яичек в горячую воду может нарушить способность к оплодотворению на последующие 6 месяцев.
№18
Наследственность и изменчивость
Генетика – это наука о законах и механизмах наследственности и изменчивости.
Задачи генетики вытекают из установленных общих закономерностей наследственности и изменчивости. К этим задачам относятся исследования:
механизмов хранения и передачи генетической информации от родительских форм к дочерним;
механизма реализации этой информации в виде признаков и свойств организмов в процессе их индивидуального развития под контролем генов и влиянием условий внешней среды;
типов, причин и механизмов изменчивости всех живых существ;
взаимосвязи процессов наследственности, изменчивости и отбора как движущих факторов эволюции органического мира.
Генетика является также основой для решения ряда важнейших практических задач. К ним относятся:
выбор наиболее эффективных типов гибридизации и способов отбора;
управление развитием наследственных признаков с целью получения наиболее значимых для человека результатов;
искусственное получение наследственно измененных форм живых организмов;
разработка мероприятий по защите живой природы от вредных мутагенных воздействий различных факторов внешней среды и методов борьбы с наследственными болезнями человека, вредителями сельскохозяйственных растений и животных;
разработка методов генетической инженерии с целью получения высокоэффективных продуцентов биологически активных соединений, а также для создания принципиально новых технологий в селекции микроорганизмов, растений и животных.
Наследственность — это неотъемлемое свойство всех живых существ сохранять и передавать в ряду поколений характерные для вида или популяции особенности строения, функционирования и развития. Наследственность обеспечивает постоянство и многообразие форм жизни и лежит в основе передачи наследственных задатков, ответственных за формирование признаков и свойств организма. Благодаря наследственности некоторые виды (например, кистеперая рыба латимерия, жившая в девонском периоде) оставались почти неизменными на протяжении сотен миллионов лет, воспроизводя за это время огромное количество поколений.
В то же время в природе существуют различия между особями как разных видов, так и одного и того же вида, сорта, породы и т. д. Это свидетельствует о том, что наследственность неразрывно связана с изменчивостью.
Изменчивость — способность организмов в процессе онтогенеза приобретать новые признаки и терять старые. Изменчивость выражается в том, что в любом поколении отдельные
особи чем-то отличаются и друг от друга, и от своих родителей. Причиной этого является то, что признаки и свойства любого организма есть результат взаимодействия двух факторов: наследственной информации, полученной от родителей, и конкретных условий внешней среды, в которых шло индивидуальное развитие каждой особи. Поскольку условия среды никогда не бывают одинаковыми даже для особей одного вида или сорта (породы), становится понятным, почему организмы, имеющие одинаковые генотипы, часто заметно отличаются друг от друга по фенотипу, т. е. по внешним признакам.
Генотип – это совокупность всех генов организма.
Фенотип - совокупность всех признаков и свойств организма, сформировавшихся в процессе его индивидуального развития. Складывается в результате взаимодействия наследственных свойств организма - генотипа и условий среды обитания.
Гомозиготность – это состояние следственного аппарата организма, при котором гомологичные хромосомы имеют одну и ту же форму данного гена.
Гетерозиготность – это наличие у диплоидного организма (или клетки) двух разных аллелей по данному гену.
Гемизиготность – это состояние, связанное с тем, что у организма один или несколько генов не парные, т. е. не имеют аллельных партнёров. Например гены половых хромосом XX и XY.
Закономерности наследования признаков установил Грегор Мендель (1865). Свои опыты по скрещиванию он проводил на горохе. Горох – самоопыляемое растение. В эксперименте были использованы чистые линии гороха, не дающие расщепления по изучаемым признакам, имеющие максимально однородную наследственность. Скрещивание двух организмов называют гибридизацией. Потомство от скрещивания двух особей с различной наследственностью называют гибридным, а отдельную особь - гибридом (от лат. hibrida - помесь).
Первый закон Менделя: