

Молекулярная биология вируса гепатита в
ВГВ был открыт во второй половине ХХ века в результате открытия так называемого австралийского антигена у аборигенов в Австралии [Blumberg B. et al., 1965]; этот антиген оказался поверхностным антигеном вируса гепатита В (HBs Ag). Вскоре, в 1970 г., Д. Дейн методом электронной микроскопии обнаружил в крови и в ткани печени больного желтухой вирус, представленный сферическими и полигональными образованиями (их назвали частицами Дейна), обладающими инфекциозностью и разнообразной антигенностью.
Вирус гепатита В (ВГВ) принадлежит к семейству Hepadnaviridae, роду Orthohepadnaviruses, представители которого инфицируют человека, человекообразных обезьян, низших приматов и грызунов. В это же семейство входят представители рода Avihepadnaviruses, инфицирующие птиц [Kidd-Ljunggren K. et al., 2002].
Гепаднавирусы преимущественно инфицируют клетки печени, но небольшое количество ДНК вирусов может быть найдено в почках, поджелудочной железе и в мононуклеарных клетках. Тем не менее, инфекция в этих местах не связана с внепеченочными заболеваниями [Ganem D., Prince A.M., 2004].
Организация генома
Вирионы состоят из липидной оболочки, в которой находятся три поверхностных гликопротеина большой (LHBsAg или пре-S1), средний (MHBsAg или пре-S2) и малый (SHBsAg или S), и внутреннего нуклеокапсида диаметром 30-34 нм. Нуклеокапсид содержит антиген сердцевины или коровый белок (HBcAg), а также вирусный геном и ДНК-полимеразу (рисунок 1б).
Геном ВГВ состоит из кольцевой частично двухцепочечной ДНК размером около 3200 нуклеотидов и полимеразы (P), которая отвечает за синтез вирусной ДНК в инфицированных клетках. На 5'-конце минус-цепь ДНК ковалентно связана с белком P. Плюс-цепь более короткая, по сравнению с минус-цепью, и 5'-конец ее фиксирован в постоянной позиции (в 200 нуклеотидах от 5'-конца минус-цепи), в то время как положение 3'-конца может варьироваться (рисунок 1а).
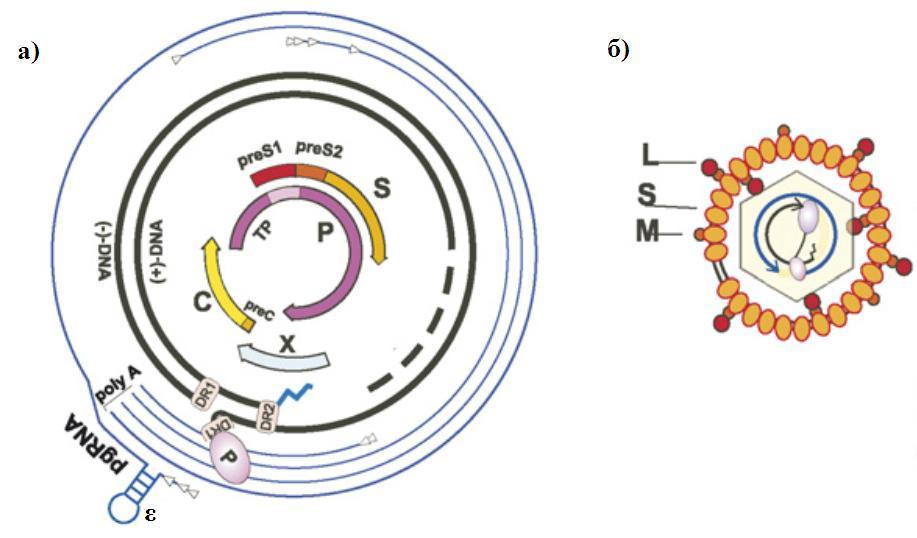
Рисунок 1. (а) Схема организации генома ВГВ. Геномная ДНК и четыре ОРС показаны в центре рисунка. Внешние линии - геномная и субгеномные РНК. Стрелками обозначены сайты начала транскрипции. ε – энкапсидационный сигнал. DR1 и DR2 – короткие повторы, размером 11п.н., которые выступают в качестве сайтов инициации синтеза полноразмерной минус-цепи и короткой плюс-цепи, соответственно. PREα и PREβ – посттранскрипционный регуляторный сигнал. (б) Схема строения ВГВ. Нужно отметить, что preS-домен белка L может быть обращен как наружу, так и внутрь оболочки. (Nassal M, 2008).
Геном ВГВ имеет лишь четыре перекрывающиеся открытые рамки считывания (ОРС), причем 80% генома занимает полимераза, которая транслируется с прегеномной РНК, с нее же транслируется и HBcAg (рисунок 2). Предшественник НВеAg – прекоровый белок, экспрессируется с другой мРНК, точка инициации которой расположена в 29-35 п.о. против хода транскрипции от точки инициации прегеномной РНК. Транскрипцию обеих этих РНК регулирует коровый промотер, который располагается вверх от С гена. Белки оболочки транслируются с двух мРНК: большой транслируется с preS1 мРНК размером 2.4 т.п.о., а средний и малый – с preS2/S мРНК размером 2.1 т.п.о. Транскрипцию этих мРНК регулирует соответственно preS1- и preS2/S-промотеры. Синтез белка Х происходит с мРНК размером 0.9 т.п.о. [Ganem D. and Varmus H., 1987]. Помимо четырех промоторов экспрессию генов регулирует 2 энхансера: ENI регулирует транскрипцию прегеномной РНК, гена S и гена X [Zhang P. et al, 1992; Su H. and Yee J.-K., 1992]. ENII регулирует транскрипцию прегеномной РНК и прекоровой РНК [Yuh C.H. and Ting L.P., 1990; Wu X. et al., 1992; Park G.T. et al., 1997].

Рисунок 2. Схема транскрипции и трансляции генома ВГВ. Один транскрипт размером 3.5 т.п.о. выступает и в качестве прегенома и в как мРНК для синтеза корового белка.
Транскрипция всех генов происходит в одном направлении и только с минус-цепи ДНК ВГВ и терминируется в общем для всех РНК поли(А)-сайте. Интересно отметить, что т.к. коровый промотор находится менее чем в 200 п.о. от этого поли(А)-сайта, поэтому сигнал полиаденилирования узнается лишь во второй раз [Guo W.T. et al., 1991].
Вирусные белки
Белки оболочки
Как уже упоминалось выше оболочка вириона состоит из трех поверхностных белков(LHBsAg или пре-S1; MHBsAg или пре-S2; SHBsAg или S), которые кодируются одной ОРС с общим стоп-кодоном (ТТА) и тремя отельными сайтами начала трансляции (ATG). HBsAg один из самых наиболее часто применяемых маркеров ВГВ, поскольку его выявление в сыворотке крови пациентов не только опережает появление клинических симптомов, но и является индикатором стадии ВГВ-инфекции.
Кроме того, HBsAg имеет важнейшее значение, так как содержит основные нейтрализующие эпитопы и поэтому используется в коммерческих вакцинах против гепатита В.
L и M белки - это дополнительные поверхностные антигены, экспрессируемые в гораздо меньшем количестве, чем HBsAg. И это несмотря на то, что включают большое количество Т и В-клеточных эпитопов. Белок L, благодаря двоякой направленности preS-домена, играет ключевую роль в связывании вируса с рецепторами клетки-хозяина, в сборке вириона и го освобождении из клетки [Bruss V. et al., 1994; Clayton R. et al., 2001; Lambert C. and Prange R., 2001].