

Вопрос 2:
РЕПЛИКАЦИЯ: синтез ДНК
Протекает в ядре в S-фазу клеточного цикла перед митозом
Стимулы: гормоны, ростовые факторы, белки-циклины
Матрица: обе нити ДНК, образуются 2 репликативные вилки
Направление синтеза новых цепей: 5′ - 3′ по принципу комплиментарности и антипараллельности
Участки синтеза – ориджины репликации
Участок ДНК между соседними ориджинами - репликон
Этапы репликации: инициация, элонгация, терминация
Субстраты и источники энергии: дАТФ, дГТФ, дТТФ, дЦТФ
Кофактор: Mg2+
Полуконсервативный процесс синтеза: каждая дочерняя молекула ДНК содержит одну родительскую нить и одну синтезированную
Образуется идентичная молекула ДНК (клетка 4n)
1 Этап репликации: инициация
Формирование репликативной вилки:
1. ДНК-топоизомераза гидролизует 3′,5′-фосфодиэфирную
связь в одной из цепей ДНК и присоединяется к 5′-концу в
точке разрыва
2. ДНК-хеликаза, используя энергию АТФ, разрывает
водородные связи и обеспечивает локальное разделение двойной
спирали ДНК
• ДНК-топоизомераза восстанавливает 3′,5′-фосфодиэфирную
связь и отделяется
• SSB (single strand binding)–белки связываются с
одноцепочечными участками, препятствуя комплементарному
скручиванию цепей
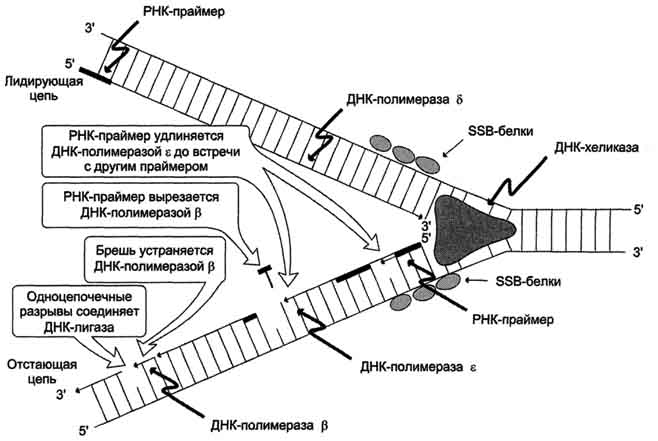
2 Этап репликации: элонгация
Синтез новых цепей ДНК
-Лидирующая цепь: 3′ - 5′ (синтез непрерывный по ходу
движения репликативной вилки)
- Отстающая цепь: 5′ - 3′ (рост этой цепи начинается после того,
как на лидирующей цепи синтезируется участок из ≈200
нуклеотидов, синтез идет против движения репликативной вилки
в виде фрагментов Оказаки)
- Синтез цепей начинается с образования «затравки» (РНК-
праймера из ≈10 нуклеотидов)
- Ферменты:
ДНК-полимераза α синтезирует РНК-праймер и небольшой
участок ДНК
ДНК-полимераза δ удлиняет лидирующую цепь
ДНК-полимераза δ или ε удлиняют отстающую цепь 3 этап репликации: терминаци
3 Этап репликации: терминация
Исключение праймеров
Завершение формирования отстающей цепи ДНК
-Эндонуклеаза (РНКаза) удаляет РНК-праймер
- ДНК-полимераза β заполняет «брешь»
- ДНК-лигаза объединяет фрагменты, затрачивая
энергию АТФ
Билет № 23
1. Молекулярная организация биологических мембран. Трансмембранный перенос веществ.
2. Структурная организация монооксигеназной системы окисления. Семейства цитохрома р450. Основные реакции, катализируемые изоферментами цитохрома р450.
Вопрос 1:
Строение и состав мембран
Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы "растворены" в липидном бислое .
Мембранные липиды - амфифильные (амфипатические) молекулы, т.е. в молекуле есть как гидрофильные группы (полярные "головки"), так и алифатические радикалы (гидрофобные "хвосты"), самопроизвольно формирующие бислой. В большинстве эукариотических клеток они составляют около 30-70% массы мембраны. В мембранах присутствуют липиды трёх главных типов - фосфолипиды, гликолипиды и холестерол (холестерин).
Липидный состав мембран различен, содержание того или другого липида, по-видимому, определяется разнообразием функций, выполняемых этими липидами в мембранах.
Фосфолипиды. Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран - фосфатидилхолины и фосфатидилэтаноламины. В мембранах эукариотических клеток обнаружено огромное количество разных фосфолипидов, причём они распределены неравномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных "головок" ,так и ацильных остатков .
Каждый глицерофосфолипид, например фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирно-кислотных остатков.
На долю глицерофосфолипидов (полярная группа - инозитол) приходится лишь 2-8% всех фосфолипидов, содержащихся в клеточной мембране эукариотов. Инозитол в составе фосфати-дилинозитолов может быть фосфорилирован по С4 (фосфатидилинозитол-4-монофосфат) или С4 и С5(фосфатидилинозитол-4,5-бисфосфат).
В состав фосфатидилинозитол-4,5-бисфосфатов входят в основном ацильные остатки стеариновой или пальмитиновой (по первому положению глицерола) и арахидоновой (по второму положению) жирных кислот.
Специфические фосфолипиды внутренней мембраны митохондрий - кардиолипины (дифосфатидилглицеролы), построенные на основе глицерола и двух остатков фосфатидной кислоты. Они синтезируются ферментами внутренней мембраны митохондрий и составляют около 22% от всех фосфолипидов мембраны.
В плазматических мембранах клеток в значительных количествах содержатся сфингомиелины .Сфингомиелины построены на основе церамида - ацилированного аминоспирта сфингозина. Полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Сфингомиелины - главные липиды миелиновой оболочки нервных волокон.
Гликолипиды. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа - углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у первого углеродного атома церамида. В зависимости от длины и строения углеводной части различают цереброзиды, содержащие моно- или олигосахаридный остаток, и ганглиозиды, к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-ацетилнейраминовую кислоту (NANA).
Полярные "головки" гликосфинголипидов находятся на наружной поверхности плазматических мембран. В значительных количествах гликолипиды содержатся в мембранах клеток мозга, эритроцитов, эпителиальных клеток. Ганглиозиды эритроцитов разных индивидуумов различаются строением олигосахаридных цепей, проявляющих антигенные свойства.
Холестерол. Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жёсткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является "полярной головкой" .
Для животной клетки среднее молярное отношение холестерол/фосфолипиды равно 0,3-0,4, но в плазматической мембране это соотношение гораздо выше (0,8-0,9). Наличие холестерола в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липи-дов и белков, и поэтому может влиять на функции: мембранных белков.
В составе мембран растений холестерола нет, а присутствуют растительные стероиды - ситостерол и стигмастерол.