

Синтез белка
Синтез белка (трансляция) является самым сложным из биосинтетических процессов. Сам процесс протекает в пять этапов.
Активация аминокислот. Каждая из 20 аминокислот белка соединяется ковалентными связями к определённой т-РНК, используя энергию АТФ. Реакция катализуется специализированными ферментами, требующими присутствия ионов магния.
Инициация белковой цепи. и-РНК, содержащая информацию о данном белке, связывается с малой частицей рибосомы и с инициирующей аминокислотой, прикреплённой к соответствующей т-РНК. т-РНК комплементарна с находящимся в составе и-РНК триплетом, сигнализирующим о начале белковой цепи.
Элонгация. Полипептидная цепь удлиняется за счёт последовательного присоединения аминокислот, каждая из которых доставляется к рибосоме и встраивается в определённое положение при помощи соответствующей т-РНК. В настоящее время генетический код полностью расшифрован, то есть всем аминокислотам поставлены в соответствие триплеты нуклеотидов. Элонгация осуществляется при помощи белков цитозоля (так называемые факторы элонгации).
Терминация. После завершения синтеза цепи, о чём сигнализирует ещё один специальный кодон и-РНК, полипептид высвобождается из рибосомы.
Сворачивание и процессинг. Чтобы принять обычную форму, белок должен свернуться, образуя при этом определённую пространственную конфигурацию. До или после сворачивания полипептид может претерпевать процессинг, осуществляющийся ферментами и заключающийся в удалении лишних аминокислот, присоединении фосфатных, метильных и других групп и т. п.
Механизмы реализации генетической информации. Генетическая информация, записанная в виде определенной последовательности нуклеотидов молекулы ДНК, обеспечивает синтез определенного белка-фермента, который катализирует течение соответствующей биохимической реакции, в результате чего проявляется признак. Геномный уровень организации генетического материала обеспечивает взаимодействие аллельных и неаллельных генов. Следовательно, проявление действия конкретного гена зависит от других генов. Они могут влиять непосредственно на данный ген, через взаимодействие
белков-ферментов, кодируемых этими генами, изменять течение биохимических реакций и, тем самым, влиять на проявление данного признака. В свою очередь данный ген может влиять на реализацию действия других генов. На реализацию действия гена влияют факторы внешней среды, которые могут изменять структуру молекул ДНК, и-РНК, белков-ферментов, течение биохимических реакций и, следовательно, - фенотипические проявления генов.
Генетический код
Генетический код обладает рядом особенностей.
Во-первых, в коде отсутствуют «знаки препинания», то есть сигналы, показывающие начало и конец кодонов.
Во-вторых, 3 нуклеотидных триплета (УАГ, УАА, УГА) не соответствуют никакой аминокислоте, а обозначают конец полипептидной цепи, а кодон АУГ сигнализирует о начале цепи либо (если он в середине последовательности) об аминокислоте метионине. Многие аминокислоты могут кодироваться несколькими различными кодонами. Все кодоны аминокислот одинаковы у всех изученных организмов: от вируса до человека.
Свойства
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин).
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
Синтез белка требует больших затрат энергии – 24,2 ккал/моль. После окончания синтеза белок при помощи специального полипептидного лидера доставляется к месту своего назначения.
Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы).
Определение кариотипа
Внешний вид хромосом существенно меняется в течение клеточного цикла: в течение интерфазы хромосомы локализованы в ядре, как правило, деспирализованы и труднодоступны для наблюдения, поэтому для определения кариотипа используются клетки в одной из стадий их деления — метафазе митоза.
Процедура определения кариотипа
Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток, для определения человеческого кариотипа используется либо одноядерные лейкоциты, извлечённые из пробы крови, деление которых провоцируется добавлением митогенов, либо культуры клеток, интенсивно делящихся в норме (фибробласты кожи, клетки костного мозга). Обогащение популяции клеточной культуры производится остановкой деления клеток на стадии метафазы митоза добавлением колхицина — алкалоида, блокирующего образование микротрубочек и «растягивание» хромосом к полюсам деления клетки и препятствующего тем самым завершению митоза.
Полученные клетки в стадии метафазы фиксируются, окрашиваются и фотографируются под микроскопом; из набора получившихся фотографий формируются т. н. систематизированный кариотип — нумерованный набор пар гомологичных хромосом (аутосом), изображения хромосом при этом ориентируются вертикально короткими плечами вверх, их нумерация производится в порядке убывания размеров, пара половых хромосом помещается в конец набора.
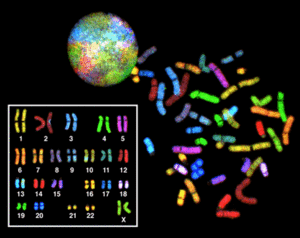
Номенклатура
Запись имеет следующий формат:
[номер хромосомы] [плечо] [номер участка].[номер полосы]
длинное плечо хромосомы обозначают буквой q, короткое — буквой p, хромосомные аберрации обозначаются дополнительными символами.
Таким образом, 2-я полоса 15-го участка короткого плеча 5-й хромосомы записывается как 5p15.2.
Для кариотипа используется запись, имеющая следующий формат:
[количество хромосом], [половые хромосомы], [особенности][5].
Для обозначения половых хромосом у различных видов используются различные символы (буквы), зависящие от специфики определения пола таксона (различные системы половых хромосом).
Половой хроматин — особые хроматиновые тельца клеточных ядер особей женского пола у человека и других млекопитающих. Располагаются у ядерной оболочки, на препаратах имеют обычно треугольную или овальную форму; размер 0,7—1,2 мк . Половой хроматин образован одной из Х-хромосом женского кариотипа и может быть выявлен в любой ткани человека (в клетках слизистых оболочек, кожи, крови, биопсированной ткани), Наиболее простым исследованием полового хроматина является исследование его в клетках эпителия слизистой оболочки полости рта. Взятый шпателем соскоб со слизистой оболочки щеки помещают на предметное стекло, окрашивают ацетоорсеином и анализируют под микроскопом 100 светлоокрашенных клеточных ядер, подсчитывая, сколько из них содержат половой хроматин. В норме он встречается в среднем в 30—40% ядер у женщин и не обнаруживается у мужчин.
В условиях патологии могут изменяться размеры телец полового хроматина, а также их число в каждом отдельном ядре и в среднем на 100 ядер. Половой хроматин изучают при цитологическом определении пола (например, при гермафродитизме); для выявления хромосомных болезней (синдром Шерешевского — Тернера, для которого характерно отсутствие полового хроматина у женщин; синдром Клайнфелтера, при котором у мужчин выявляют половой хроматин; синдром трисомии X, при котором в ядре вместо одного тельца полового хроматина выявляют два); при ряде патологических процессов, особенно злокачественных (в частности, для решения вопроса о виде гормональной терапии при раке молочной железы); для характеристики действия ряда фармакологических средств (например, кортикостероидов — по изменению ими количества клеток, содержащих половой хроматин) и др.