

kossitsky
.pdfЧАСТНАЯ ФИЗИОЛОГИЯ АНАЛИЗАТОРОВ
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Зрительный анализатор (или зрительная сенсорная система) — важнейший из органов чувств человека и большинства высших позвоночных животных. Он дает более 90% информации, идущей к мозгу от всех рецепторов (не случайна пословица «Лучше один раз увидеть, чем сто раз услышать»). Благодаря опережающему эволюционному развитию именно зрительных механизмов мозг хищных животных и приматов претерпел резкие изменения и достиг значительного совершенства. Зрительное восприятие — многозвеньевой процесс, начинающийся с проекции изображения на сетчатку глаза и возбуждения фоторецепторов и заканчивающийся принятием высшими отделами зрительного анализатора, локализованными в коре мозга, решения о наличии в поле зрения того или иного зрительного образа.
Оптическая система глаза. В связи с необходимостью наводить глаз на рассматриваемый объект, вращая его, природа создала у большинства видов животных шарообразную форму глазного яблока. На пути к светочувствительной оболочке глаза — сетчатке лучи света проходят через несколько прозрачных поверхностей—переднюю и заднюю поверхности роговицы, хрусталика и стекловидного тела. Разная кривизна и показатели преломления этих поверхностей определяют преломление световых лучей внутри глаза.
Преломляющую силу любой оптической системы выражают в диоптриях (D). Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила глаза человека составляет 59 D при рассматривании далеких и 70,5 D при рассматривании близких предметов.
Сложность оптической системы глаза затрудняет точную оценку хода лучей внутри него и оценку размера изображения на сетчатке. Поэтому пользуются упрощенной моделью — «редуцированным глазом», в котором все преломляющие среды имеют один и тот же показатель преломления в единую сферическую поверхность. Для того чтобы построить на сетчатке такого схематического глаза изображение, нужно провести линии от концов рассматриваемого предмета к узловой точке, лежащей в 7,15-10-3 м (7,15 мм) кзади от роговой оболочки и на 15 мм (15-10-3 м) вперед от сетчатки. При этом изображение на сетчатке получается действительным, уменьшенным и обратным (рис. 210). Для построения изображения на сетчатке нужно знать величину предмета и его расстояние от роговицы глаза. Из подобия треугольника АВО и аbО на рис. 210 ясно, что
 т. е. изображение на сетчатке (ab) во столько раз меньше самого предме-
т. е. изображение на сетчатке (ab) во столько раз меньше самого предме-
та ( А В ) , во сколько dO (1 5 мм) меньше расстояния предмета от глаза (DO).
Рис. 210. Ход лучей и построение изображения в редуцированном глазу.
АВ — предмет; ab — его изображение; О — узловая точка; D — d — главная оптическая ось.
441

Аккомодация
Для ясного видения предмета необходимо, чтобы лучи от его точек попадали на поверхность сетчатки, т. е. были здесьсфокусированы (рис. 211). Когда человек смотрит на далекие предметы, их изображение сфокусировано на сетчатке, и они видны ясно. При этом близкие предметы видны неясно, их изображение на сетчатке расплывчато, так как лучи от них собираются за сетчаткой. Видеть одновременно одинаково ясно предметы, удаленные от глаза на разное расстояние, невозможно. В этом легко убедиться: переводя взгляд с близкого на далекие предметы, вы перестаете его ясно видеть.
Приспособление глаза к ясному видению удаленных на разное расстояние предметов называется аккомодацией (аккомодацию глаза не следует смешивать с аккомодацией возбудимой ткани). При аккомодации происходит изменение кривизны хрусталика и, следовательно, его преломляющей способности. При рассматривании близких предметов хрусталик делается более выпуклым (рис. 212), благодаря чему лучи, расходящиеся от светящейся точки, сходятся на сетчатке.
Радиус кривизны передней поверхности хрусталика при аккомодации уменьшается с 10 до 6 мм (с 1-10-2 м до 6-10-3 м), а задней поверхности с 6 до 5,5 мм (с 6-10-3 м до 5,5-10-3 м).
Механизм аккомодации сводится к сокращению ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключен в тонкую прозрачную капсулу, переходящую по краям в волокна цинновой связки, прикрепленной к ресничному телу. Эти волокна всегда натянуты и растягивают капсулу, сжимающую и уплощающую хрусталик. В ресничном теле находятся гладкомышечные волокна. При их сокращении тяга
Рис. 212. Механизм аккомодации (по Гельмгольцу).
В левой половине хрусталик (7) уплощен при рассматривании далекого предмета, а справа он стал более выпуклым за счет аккомодационного усилия при рассматривании близкого предмета. 1 — склера; 2 — сосудистая оболочка; 3 — сетчатка; 4 — роговица; 5 — передняя камера; 6 — радужная оболочка; 7 — хрусталик; 8 — стекловидное тело; 9 — ресничная мышца, ресничные отростки и ресничные связки (ц инновы); 10 — центральная ямка; II —зрительный нерв.
цинновых связок ослабляется, а значит уменьшается давление на хрусталик, который вследствие своей эластичности принимает более выпуклую форму. Таким образом, ресничные мышцы являются аккомодационными мышцами. Они иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, и, следовательно, ограничивает аккомодацию глаз при рассмотрении близких предметов. Наоборот, парасимпатомиметические вещества — пилокарпин и эзерин — вызывают сокращение этой мышцы.
Для нормального глаза молодого человека дальняя точка ясного видения лежит в бесконечности. Далекие предметы он рассматривает без всякого напряжения аккомодации, т. е. без сокращения ресничной мышцы. Ближайщая точка ясного видения находится на расстоянии 10 см от глаза. Предметы, расположенные ближе 10 см, не могут быть ясно видны человеком с нормальным зрением даже при максимальном сокращении ресничной мышцы, т. е. при максимальном аккомодационном усилии. Сила аккомодации может быть выражена в диоптриях.
Если ближайшая точка ясного видения находится на расстоянии 10 см от глаза человека, то линза с фокусным расстоянием в 10 см, т. е. в 10 D, превратит лучи, идущие от ближайшей ясно видимой точки, в параллельные. Следовательно, при помощи линзы устранится необходимость в аккомодации. Поэтому можно заменить максимальное аккомодационное усилие человека, поместив у него перед глазом линзу силой в 10 D, отсюда следует, что максимальная сила аккомодации равна 10 D.
Старческая дальнозоркость. Хрусталик с возрастом становится менее эластичным и при ослаблении натяжения цинновых связок его выпуклость или не изменяется, или увеличивается лишь незначительно. Поэтому ближайшая точка ясного видения отодвигается от глаз. Это состояние называется старческой дальнозоркостью, или пресбиопией. Поэтому пожилые люди исправляют недостаток аккомодации с помощью двояковыпуклых линз.
Аномалии рефракции глаза
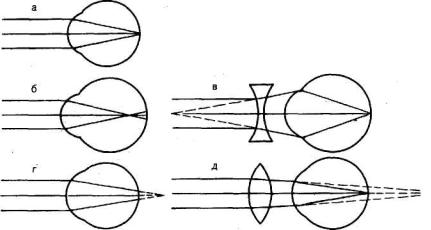
Существуют две главные аномалии преломления лучей (рефракции) в глазу: близорукость, или миопия, и дальнозоркость, или гиперметропия. Эти аномалии обусловлены, как правило, не недостаточностью преломляющих сред, а ненормальной длиной глазного яблока (рис. 213).
Близорукость. Если продольная ось глаза слишком длинная, то главный фокус будет находиться не на сетчатке, а перед ней, в стекловидном теле. В этом случае параллельные
Рис. 213. Схема рефракции в нормальном ( а ), близоруком (б) и дальнозорком (г) глазу. Оптическая коррекция близорукости ( в ) и дальнозоркости (д).
443
лучи сходятся в одну точку не на сетчатке, а где-то ближе нее, а на сетчатке вместо точки возникает круг светорассеяния. Такой глаз называется близоруким — миопическим. У близорукого дальняя точка ясного видения находится не в бесконечности, а на конечном (и довольно близком) расстоянии. Чтобы ясно видеть вдаль, близорукий должен поместить перед глазами вогнутые стекла, которые уменьшат преломляющую силу хрусталика и тем самым отодвинут сфокусированное изображение на сетчатку.
Дальнозоркость. Противоположностью близорукости является дальнозоркость — гиперметропия. В дальнозорком глазу продольная ось глаза короткая, и поэтому параллельные лучи, идущие от далеких предметов, собираются сзади сетчатки, а на ней получается неясное, расплывчатое изображение предмета. Этот недостаток рефракции может быть компенсирован путем аккомодационного усилия, т. е. увеличения выпуклости хрусталика. Поэтому дальнозоркий человек напрягает аккомодационную мышцу, смотря не только вблизи, но и вдаль.
У дальнозорких людей ближайшая точка ясного видения отстоит от глаза дальше, чем у лиц с нормальным зрением. Поэтому аккомодационные усилия при рассматривании близких предметов являются недостаточными. В результате для чтения дальнозоркие люди должны надевать двояковыпуклые очки, усиливающие преломление лучей.
Гиперметропию не следует смешивать со старческой дальнозоркостью. Эти два недостатка имеют общим только то, что при них необходимо пользоваться двояковыпуклыми очками.
Астигматизм. К аномалиям рефракции следует отнести также астигматизм, т. е. неодинаковое преломление лучей в разных направлениях (например, по горизонтальному и вертикальному меридиану). Все люди в небольшой степени являются астигматиками, поэтому астигматизм следует отнести к несовершенству строения глаза как оптического инструмента.
Астигматизм обусловлен тем, что роговая оболочка не является строго сферической поверхностью: в различных направлениях она имеет различный радиус кривизны. При сильных степенях астигматизма эта поверхность приближается к цилиндрической, что дает искаженное изображение на сетчатке. Исправляется астигматизм помещением перед глазами специальных цилиндрических стекол. Если, например, роговая оболочка преломляет слабее в вертикальном направлении, то стекло должно преломлять в этом направлении сильнее.
Зрачковый рефлекс
Зрачком называется отверстие в центре радужной оболочки, через которое лучи света проходят внутрь глаза. Зрачок способствует четкости изображения предметов на сетчатке, пропуская только центральные лучи и устраняя так называемую сферическую аберрацию. Сферическая аберрация состоит в том, что лучи, попавшие на периферические части хрусталика, преломляются сильнее центральных лучей. Поэтому, если не устранять периферических лучей, на сетчатке должны получаться круги светорассеяния. Мускулатура радужной оболочки способна изменять величину зрачка и тем самым регулировать поток света, попадающий в глаз. Оптимальным в условиях дневного зрения является диаметр зрачка 2,4 мм (2,4 • 10-3 м). На ярком свету зрачок уменьшается в диаметре до 1,8 мм (1,8-10-3 м). В темноте зрачок расширяется (до 7,5 мм, т. е. 7,5-10-3 м в диаметре), что приводит к ухудшению качества изображения на сетчатке, но увеличивает световую чувствительность зрительной системы. Таким образом, изменение диаметра зрачка изменяет световой поток в 17 раз. Связь между интенсивностью освещения и диаметром зрачка логарифмическая. Реакция зрачка на изменение освещенности носит адаптивный характер, так как несколько (хотя и в небольшом диапазоне) стабилизирует уровень освещения сетчатки. Если прикрыть глаз от света, а затем открыть его, расширившийся при затемнении зрачок быстро суживается. Это сужение происходит рефлекторно («зрачковый рефлекс»).
АДА.
В радужной оболочке имеется два вида мышечных волокон, окружающих зрачок: одни — кольцевые (m.sphincter iridis), иннервируемые парасимпатическими волокнами глазодвигательного нерва, другие — радиальные (m.dilatator iridis), иннервируемые симпатическими нервами. Сокращение первых вызывает сужение зрачка, сокращение вторых — его расширение. Соответственно этому ацетилхолин и эзерин вызываютсужение, а адреналин — расширение зрачка. При эмоциях, сопровождающихся возбуждением симпатической системы (страх, ярость), а также во время боли зрачки расширяются. Поэтому расширение зрачка — важный симптом ряда патологических состояний (например, болевого шока). Зрачки расширяются также при гипоксии. Поэтому расширение зрачков при глубоком наркозе указывает нанаступающую гипоксию и является признаком опасного для жизни состояния.
Сужение зрачка наступает при рассматривании близко находящихся предметов, когда происходит аккомодация и сведение зрительных осей обоих глаз (конвергенция).
Зрачки обоих глаз у здоровых людей бывают расширены или сужены одинаково. При освещении одного глаза зрачок другого тоже суживается, такая реакция называется содружественной.
В некоторых случаях размеры зрачков обоих глаз различны (анизокория). Это может происходить вследствие поражения симпатического нерва на одной стороне, что влечет за собой сужение зрачка (миоз) и одновременно сужение глазной щели (симптом Горнера). Расширение зрачка (мидриаз) одного глаза может быть вызвано параличом n.oculomotorius или раздражением п. sympathicus.
Рецепторный аппарат зрительного анализатора. Структура и функция отдельных слоев сетчатки
Сетчатка представляет собой внутреннюю оболочку глаза, имеющую сложную многослойную структуру (рис. 214). Здесь расположены два вида различных по своему функциональному значению фоторецепторов — палочки и колбочки и несколько видов нервных клеток с их многочисленными отростками.
Под влиянием световых лучей в фоторецепторах происходят фотохимические реакции, состоящие в изменении светочувствительных зрительных пигментов. Это вызывает возбуждение фоторецепторов и затем синаптическое возбуждение связанных с палочками и колбочками нервных клеток. Последние образуют собственно нервный аппарат глаза, который передает зрительную информацию в центры головного мозга и участвует в ее анализе и переработке. Таким образом, сетчатка является как бы частью мозга, вынесенной на периферию.
Рассмотрим структуру и функцию слоев сетчатки, следуя от наружного (заднего, наиболее удаленного от зрачка) слоя сетчатки к внутреннему (расположенному ближе к зрачку) ее слою.
Пигментный слой сетчатки. Наружный слой сетчатки образован пигментным эпителием, содержащим пигмент фусцин. Этот пигмент поглощает свет, препятствуя его отражению и рассеиванию, что способствует четкости зрительного восприятия.
У некоторых ночных животных между фоторецепторами и пигментными клетками имеется отражающий свет слой, состоящий из особых кристаллов или нитей. Отражение от них света является причиной свечения глаз у ночных животных при внешнем освещении. Наличие слоя,отражающего свет, обусловливает действие на фоторецепторы не только прямых, но и отраженных лучей, что в условиях слабой освещенности повышает возможность восприятия света.
Пигментные клетки, отростки которых окружают светочувствительные членики палочек и колбочек, принимают участие в обмене веществ в фоторецепторах и в синтезе зрительного пигмента.
Фоторецепторы. К слою пигментного эпителия изнутри примыкает слой фоторецепторов, которые своими светочувствительными члениками обращены в сторону, противоположную свету.
445
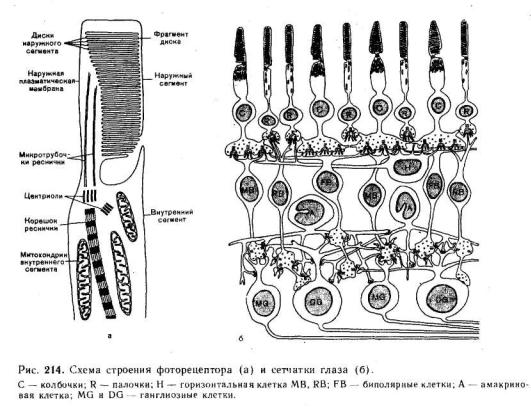
Каждый фоторецептор — палочка или колбочка — состоит из чувствительного к действию света наружного сегмента, содержащего зрительный пигмент, и внутреннего сегмента, содержащего ядро и митохондрии, обеспечивающие энергетические процессы в фоторецепторной клетке.
Электронно-микроскопические исследования выявили, что наружный сегмент каждой палочки состоит из 400—800 тонких пластинок, или дисков, диаметром около 6 мкм (6-10-6 м). Каждый диск представляет собой двойную мембрану, состоящую из мономолекулярных слоев липидов, находящихся между слоями молекул белка. С молекулами белка связан ретиналь, входящий в состав зрительного пигмента родопсина.
Наружный и внутренний сегменты фоторецепторной клетки разделены мембранами, через которые проходит пучок из 16—18 тонких фибрилл. Внутренний сегмент переходит в отросток, с помощью которого фоторецепторная клетка передает возбуждение через синапс на контактирующую с ней биполярную нервную клетку.
Учеловека в глазу имеется около 6—7 млн. колбочек и 110—125 млн. палочек. Палочки
иколбочки распределены в сетчатке неравномерно. Центральная ямка сетчатки (fovea centralis) содержит только колбочки (до 140 000 колбочек на 1 мм2). По направлению к периферии сетчатки число колбочек уменьшается, а количество палочек возрастает.
Периферия сетчатки содержит почти исключительно палочки. Колбочки функционируют в условиях яркой освещенности и воспринимают цвета; палочки являются рецепторами, воспринимающими световые лучи в условиях сумеречного зрения.
Уночных животных, например у совы и летучей мыши, в сетчатке преобладают палочки,
удневных животных — голубей, кур, ящериц — колбочки. У животных, ведущих смешанный образ жизни (хищные, приматы), в сетчатке есть оба вида рецепторов.
Раздражение различных участков сетчатки показывает, что различные цвета воспринимаются лучше всего при действии световых раздражителей на центральную ямку, где
446

расположены почти исключительно колбочки. По мере удаления от центра сетчатки восприятие цвета становится все хуже. Периферия сетчатки, где находятся исключительно палочки, не воспринимает цвета. Световая чувствительность колбочкового аппарата сетчатки во много раз меньше таковой элементов, связанных с палочками. Поэтому в сумерках, в условиях малой освещенности, центральное колбочковое зрение резко понижено и преобладает периферическое палочковое зрение. Так как палочки не воспринимают цветов, то в сумерках человек цвета не различает.
Нарушение функций палочек, возникающее, например, при недостатке в пище витамина А, вызывает расстройство сумеречного зрения, так называемую куриную слепоту: человек совершенно слепнет в сумерках, но днем зрение остается нормальным. Наоборот, при поражении колбочек возникает светобоязнь: человек видит только при слабом свете и слепнет при ярком освещении. В этом случае может развиться и полная слепота на цвета — ахромазия.
Слепое пятно. Место входа зрительного нерва в глазное яблоко — сосок зрительного нерва — не содержит фоторецепторов и поэтому нечувствительно к свету; это так называемое слепое пятно. В существовании слепого пятна можно убедиться с помощью опыта Мариотта. Если, закрыв левый глаз, правым фиксировать крест, изображенный на рис. 215, то при определенном расстоянии рисунка от глаз (от 10 до 25 см, т. е. от 0,1 до 0,25 м) круг исчезает, так как его изображение падает на слепое пятно.
Нейроны сетчатки. Кнутри от слоя фоторецепторных клеток в сетчатке расположен слой биполярных нейронов, к которым изнутри примыкает слой ганглиозных нервных клеток.
Аксоны ганглиозных клеток образуют волокна зрительного нерва. Таким образом, возбуждение, возникающее в фоторецепторе при действии света, передается на волокна зрительного нерва через нервные клетки — биполярные и ганглиозные.
В синапсах между биполярными и ганглиозными клетками выявлена холинэстераза; это служит указанием на то, что передача импульса с одной клетки на другую совершается с помощью медиатора ацетилхолина.
На 130 млн. фоторецепторных клеток приходится всего около 1 млн. 250 тыс. волокон зрительного нерва, являющихся отростками ганглиозных клеток. Это значит, что импульсы от многих фоторецепторов конвергируют к одной ганглиозной клетке. Один биполярный нейрон связан со многими палочками и несколькими колбочками, а одна ганглиозная клетка в свою очередь связана со многими биполярными клетками. Таким образом, каждая ганглиозная клетка суммирует возбуждение, возникающее в большом числе фоторецепторов. Лишь в центре сетчатки, в районе центральной ямки, каждая колбочка соединена с одной так называемой карликовой биполярной клеткой, с которой соединена также всего одна ганглиозная клетка.
447
Фоторецепторы, соединенные с одной ганглиозной клеткой, образуют рецептивное поле ганглиозной клетки. Рецептивные поля различных ганглиозных клеток частично перекрывают друг друга. Взаимодействие соседних нейронов сетчатки обеспечивается так называемыми горизонтальными и амакриновыми клетками, отростки которых соединяют по горизонтали биполярные и ганглиозные клетки. Амакриновые клетки осуществляют процесс горизонтального, или бокового торможения между соседними элементами.
Кроме афферентных волокон, в сетчатке имеются и центробежные, или эфферентные, нервные волокна, приносящие к ней импульсы из ЦНС. Полагают, что эти импульсы действуют на синапсы между биполярными и ганглиозными клетками сетчатки и тем самым регулируют проведение возбуждения между ними. Второй тип центробежных нервных волокон представляет собой сосудодвигательные волокна, изменяющие кровоснабжение сетчатки.
Фотохимические реакции в рецепторах сетчатки
Еще в конце 70-годов прошлого столетия в сетчатке глаза животных были открыты светочувствительные пигменты, которые на свету обесцвечиваются. В палочках сетчатки человека и многих животных содержится пигмент родопсин, или зрительный пурпур, состав, свойства и химические превращения которого подробно изучены в последние десятилетия. В колбочках найден пигмент йодопсин. В колбочках имеются также пигменты хлоролаб и эритролаб; первый из них поглощает лучи, соответствующие зеленой, а второй — красной части спектра. Имеются, возможно, и другие пигменты.
Родопсин представляет собой высокомолекулярное соединение (молекулярная масса 270 000), состоящее из ретиналя — альдегида витамина А и белка опсина. При действии кванта света происходит цикл фотофизических и фотохимических превращений этого вещества: ретиналь изомеризуется, его боковая цепь выпрямляется, связь ретиналя с белком нарушается, активируются ферментативные центры белковой молекулы. При этом вначале образуются некоторые промежуточные вещества — люмиродопсин и метародопсин, после чего ретиналь отщепляется от опсина. Под влиянием фермента, названного редуктазой ретиналя, последний переходит в витамин А.
При затемнении глаз происходит регенерация зрительного пурпура, т. е. ресинтез родопсина. Для этого процесса необходимо, чтобы сетчатка получала цис-изомер витамина А, из которого образуется ретиналь. Если же витамин А в организме отсутствует, образование родопсина резко нарушается, что и приводит к развитию упомянутой выше куриной слепоты.
Фотохимические процессы в сетчатке происходят весьма экономно, т. е. при действии даже очень яркого света расщепляется только небольшая часть имеющегося в палочках родопсина.
При действии света интенсивностью 100 лк через 5 с в каждой палочке расщепляется всего около 0,006% родопсина.
Поглощение света родопсином и его расщепление различны в зависимости от длины волны действующих на него световых лучей. Родопсин, экстрагированный из сетчатки глаза человека, обнаруживает максимальное поглощение световых лучей с длиной волны около 500 нм, т. е. в сине-зеленой части спектра. Эти лучи в темноте кажутся наиболее яркими. Сравнение поглощения родопсином света с разной длиной волны и чувствительности глаза человека в темноте к различным цветам спектра обнаруживает полное их совпадение.
Структура йодопсина близка к родопсину. Йодопсин представляет собой также соединение ретиналя с белком опсином, который образуется в колбочках и отличается от опсина палочек.
Поглощение света родопсином и йодопсином различно. Йодопсин в наибольшей степени поглощает желтый свет с длиной волны около 560 нм.
448

Электрические явления в сетчатке и зрительном нерве
Фотохимические изменения в рецепторах представляют собой начальное звено в цепи трансформации световой энергии в нервное возбуждение. Вслед за ними в рецепторах,
а затем в нейронах сетчатки генерируются Рис. 216. Электроретинограмма (по Граниту). Электрические потенциалы, Отражающие а, b, с и d — волны ЭРГ; стрелками указаны моменпараметры действующего света. ты .включенияи выключениявспышки света.
Электроретинограмма. Суммарный электрический ответ сетчатки глаза на свет носит название электроретинограммы и может быть зарегистрирован от целого глаза или же непосредственно от сетчатки. Для записи электроретинограммы один электрод помещают на поверхности роговой оболочки, а другой прикладывают к коже лица вблизи глаза или мочке уха.
На электроретинограмме большинства животных, регистрируемой при освещении глаза в течение 1—2 с, различают несколько характерных волн (рис. 216). Первая волна а представляет собой небольшое по амплитуде электроотрицательное колебание. Оно переходит в быстро нарастающую и медленно убывающую электроположительную волну b, имеющую значительно большую амплитуду. После волны b, нередко наблюдается медленная электроположительная волна с. В момент прекращения светового раздражения появляется еще одна электроположительная волна d. Электроретинограмма человека имеет аналогичную форму с тем лишь отличием, что на ней между волнами а и b отмечается кратковременная волна х.
Волна а отражает возбуждение внутренних сегментов фоторецепторов (поздний рецепторный потенциал) и горизонтальных клеток. Волна b возникает в результате активации глиальных (мюллеровских) клеток сетчатки ионами калия, выделяющимися при возбуждении биполярных и амакриновых нейронов; волна с — клеток пигментного эпителия, а волна d — горизонтальных клеток.
Амплитуда всех волн электроретинограммы увеличивается пропорционально логарифму силы света и времени, в течение которого глаз находился в темноте. Только волна d (реакция на выключение) тем больше, чем длительнее действовал свет.
Электроретинограмма хорошо отражает также такие свойства светового раздражителя, как его цвет, размер и длительность действия. Поскольку на ней в интегральном виде отражена активность практически всех клеточных элементов сетчатки (кроме ганглиозных клеток), этот показатель широко используется в клинике глазных болезней для диагностики и контроля лечения при различных заболеваниях сетчатки.
Электрическая активность путей и центров зрительного анализатора. Возбуждение ганглиозных клеток сетчатки приводит к тому, что по их аксонам — волокнам зрительного нерва — в мозг устремляются электрические сигналы. В пределах же самой сетчатки передача информации о действии света происходит безимпульсным путем (распространением и транссинаптической передачей градуальных потенциалов). Ганглиозная клетка сетчатки — это первый нейрон «классического» типа в прямой цепи передачи информации от фоторецепторов к мозгу.
Различают три основных типа ганглиозных клеток; отвечающие на включение света (оп-р.еакция), его выключение (off-реакция) и на то и другое (on-off-реакция) (рис. 217). Отведение импульсов от одиночного волокна зрительного нерва микроэлектродом при точечном световом раздражении разных участков сетчатки позволило исследовать рецептивные поля ганглиозных клеток, т. е. ту часть поля рецепторов, на стимуляцию которой нейрон отвечает импульсным разрядом. Оказалось, что в центре сетчатки рецептивные поля маленькие, а на периферии сетчатки они значительно больше по диаметру. Форма их круглая, причем построены эти поля в большей части случаев концентрически: возбудительный центр и тормозная периферическая кольцевая зона или наоборот. Центр
15 Физиологиячеловека |
449 |

рецептивного поля обладает максимальной световой чувствительностью, а на краях его чувствительность снижается. При увеличении размера светового пятнышка, вспыхивающего в пределах центра рецептивного поля, величина ответа ганглиозной клетки увеличивается (пространственная суммация). Возбуждение двумя стимулами близко расположенных ганглиозных клеток приводит к их взаимному торможению: ответы каждой клетки делаются меньше, чем при одиночном раздражении. В основе этого эффекта, обеспечиваемого амакриновыми клетками сетчатки, лежит горизонтальное или боковое торможение. Рецептивные поля соседних ганглиозных клеток частично перекрываются, так что одни и те же рецепторы могут вносить вклад в генерацию ответов нескольких нейронов. Благодаря круглой форме рецептивные поля ганглиозных клеток сетчатки производят так называемое точечное описание сетчаточного изображения: оно отображается очень тонкой мозаикой, состоящей из точек — возбужденных нейронов.
Такой же характер имеет и картина возбуждения в нейронных слоях подкоркового зрительного центра — наружного коленчатого тела, куда приходят волокна из сетчатки. Рецептивные поля этих нейронов также круглые, но меньшего размера, чем в сетчатке. Фазический характер их ответов также выражен больше, чем в сетчатке. На уровне наружного коленчатого тела происходит взаимодействие афферентных сигналов, идущих из сетчатки, с эфферентными сигналами из коры, а также из слуховой и других сенсорных
450