

Ярыгин биология книга 1 2004
.pdfИногда возникающие соматические мутации являются причиной появления злокачественных новообразований. На рис. 4.2 представлена последовательность событий, приводящих к образованию таких опу холей. Как правило, в основе лежит повреждение ДНК, вызываемое внутренними факторами (нарушением процессов репликации, репа рации или рекомбинации) или внешними воздействиями (ионизиру ющей радиацией, химическими мутагенами или вирусами). Одним из результатов такого повреждения ДНК может оказаться появление клона клеток, обладающего дефектами регуляции клеточного размно жения, что приводит к опухолевому росту.
Причиной злокачественного разрастания ткани могут быть также нарушение митоза и неравноценное распределение хромосом между дочерними клетками с возникновением анэуплоидий или хромосом ных аберраций. Это вызывает либо гибель клеток, либо приводит к появлению клонов, способных к неконтролируемому росту. В злока чественных образованиях обычно встречаются субклоны, имеющие разные кариотипы, что свидетельствует о множественных аномалиях митоза в клетках опухолей.
Так как в основе злокачественного перерождения тканей может лежать изменение наследственного материала клеток, становится оче видной важная роль мутагенных факторов в процессе возникновения опухолей. Одним из таких мутагенных факторов являются вирусы, которые могут индуцировать в хромосомах то или иное мутационное изменение. Среди опухолей человека вирусное происхождение имеет
лимфома Беркитта.
Пониманию молекулярно-биологических механизмов опухолевого роста способствовало открытие так называемых онкогенов, вызываю щих злокачественное перерождение клеток и входящих в состав генома ретровирусов. Геном ретровируса состоит из одноцепочечной РНК и содержит ген обратной транскриптазы. При проникновении вируса в клетку-хозяина под контролем фермента обратной транскриптазы образуются множественные копии генетической информации вируса, но уже в виде двухцепочечной ДНК, которые встраиваются в ДНК клетки-хозяина. Функционирование таких ДНК-копий в составе ге нома клетки приводит к синтезу вирусных РНК и белков, причем вирусный онкоген (v-onc) трансформирует клетку-хозяина в опухо левую.
Использование ДНК-зондов на основе ретровирусных онкогенов обнаружило их гомологию некоторым собственным нуклеотидным последовательностям генома клеток. Эти участки получили название
протоонкогенов или клеточных онкогенов (с-опс). Протоонкогены уча ствуют в контроде клеточного роста, но в обычном состоянии не приводят к опухолевой трансформации. Их мутантные аллели, стиму лируя митоз, могут вызвать рост опухолей. Иногда активация клеточ-
ного онкогена обусловливается единичной точковой мутацией в нем.
Вдругих случаях нет необходимости в такой мутации, так как транс формирующий эффект наблюдается при присоединении онкогена к фрагментам ДНК, обладающим сильными промоторными свойствами.
Втаких ситуациях следует допустить,' что протоонкогенам присущи свойства транспозонов, или «прыгающих генов».
Предполагают, что вирусные онкогены на каком-то этапе эволю ции произошли от клеточных онкогенов, интегрированных в геном вируса. Причины, обусловливающие большую трансформирующую активность вирусных онкогенов в сравнении с клеточными, до насто ящего времени не вполне ясны.
Вопухолевых клетках часто наблюдаются хромосомные аномалии, причем некоторые опухоли отличаются наличием специфических хро мосомных дефектов.
Установлено, что онкогены нередко обнаруживаются в непосред ственной близости от точек разрывов, происходящих при опухолеспецифичных хромосомных перестройках. Это подтверждает их роль в злокачественной трансформации клеток.
4.2.2. Генеративные мутации
Изменения наследственной программы половых клеток человека приводят к рождению потомства с различными наследственно обус ловленными болезнями, в зависимости от ранга мутаций — генными
или хромосомными.
Различные генные мутации по-разному сказываются на жизнеспо собности организма, причем в случае их рецессивности они могут долго не проявляться фенотипически у потомков. Хромосомные перестройки
игеномные мутации приводят к выраженным отклонениям в развитии
ичасто являются причиной гибели организма на разных стадиях его онтогенеза, обычно в раннем эмбриогенезе. В значительной степени именно этими мутациями определяется высокий процент (15%) пре рывания диагностированных беременностей.
Триплоидии плода, как правило, приводят к прерыванию беремен ности на ранних стадиях, однако описано очень небольшое число случаев живорождения триплоидов. Анэуплоидия по разным хромосо мам встречается как в материале абортусов, так и у рожденных детей. Некоторые анэуплоидий несовместимы с жизнью. Так, трисомия по 16-й хромосоме обнаруживается только в материале абортусов. В то же время у человека известны синдромы, связанные с аномалиями числа хромосом, характеризующиеся разной степенью жизнеспособ ности.
180 |
181 |

Й ЧЙ XX «МЬъкй л
5 6 X
К% ЛХ ДА Л<4 Л» Л/.
|
|
|
10 |
11 |
12 |
Л / \ |
/!•*» АЛ J4A |
*.*. |
|
||
13 |
14 |
15 |
16 |
17 |
|
*К |
аг» |
|
|
|
* |
18 |
19 |
20 |
22 |
21 |
Y |
Б
Рис. 4.3. Синдром трисомии 21 (синдром Дауна). А—внешний вид больного; Б- кариотип больного
Наиболее частым хромосомным |
|
||
заболеванием у человека является |
|
||
синдром Дауна, обусловленный три- |
|
||
сомией по 21-й хромосоме, встре |
|
||
чающийся с частотой 1—2 на 1000 |
|
||
(рис. 4.3). Примерно в 60% случаев |
|
||
трисомия 21 является причиной ги |
|
||
бели плода, около 30% родившихся |
|
||
умирает на первом году жизни. Еще |
|
||
46% не переживает Злетний рубеж, |
|
||
однако иногда люди с синдромом |
|
||
Дауна доживают до значительного |
|
||
возраста (рис. 4.4), хотя в целом |
|
||
продолжительность их жизни со |
|
||
кращена. Применение эффектив |
|
||
ных противомикробных препаратов |
|
||
позволяет |
несколько |
увеличить |
|
продолжительность жизни таких |
|
||
больных. Трисомия 21 может быть |
Рис. 4.4. Женщина с синдромом Дауна в |
||
результатом |
случайного нерасхож |
возрасте 38 лет |
|
дения гомологичных |
хромосом в |
|
|
мейозе. Наряду с этим известны случаи регулярной трисомии, свя занной с транслокацией 21-й хромосомы на другую — 21, 22, 13, 14 или 15-ю хромосому (рис. 4.5).
}( К И U и и *
Ц |
U |
П |
и |
n |
» |
|
|
|
|
|
12 |
н |
14 |
л/ |
16 |
17 |
|
13 |
15 |
18 |
|||
IX |
М |
ДА |
|
|
|
21 |
22 |
|
|
||
19 |
20 |
|
|
Рис. 4.5. Кариотип при транслокационном синдроме Дауна (одна 21-я хромосома присоединена к 15-й хромосоме —указано стрелкой)
183

4Я00ШШ'ШІШІ^<,
UUn ть
. 1 |
2 |
3__/ |
Ч5 |
ШШхШяяш
6-Х-12
С
АЛЛйАЛЛ XX XX «Л
|
13-15 |
, |
|
|
. 16 |
|
|
17 18 , |
|||
' |
D |
|
|
|
|
|
E |
|
: |
|
|
XXXX |
AA |
ft* |
* |
|
|||||||
|
19-20 |
|
21 |
|
|
22 . Y |
|
|
|
||
|
|
|
|
|
|
Q |
|
|
|
|
|
Б
Рис. 4.6. Синдром трисомии 13 (синдром Патау). А — внешний вид больного; Б кариотип больного с трисомией в группе D:
/—аномалии лица, //—двусторонняя полисиндактилия стоп
)гіні hn
4-Ь
UUnumnm
• |
. 6-Х-12 |
|
_ _ . |
|
|
• |
|
- V |
і |
^ |
|
|
|
||||
13-15 |
|
16 |
17 18 |
||
X X К X |
Л Л Л А |
|
|
||
19-20 |
v 21 |
22 ' |
|
|
|
Рис. 4.7. Синдром трисомии 18 (синдром Эдвардса). А —внешний вид больного; Б • кариотип больного при трисомии в группе Е:
/ — черепно-лицевые аномалии, //—характерное расположение пальцев на кистях больного
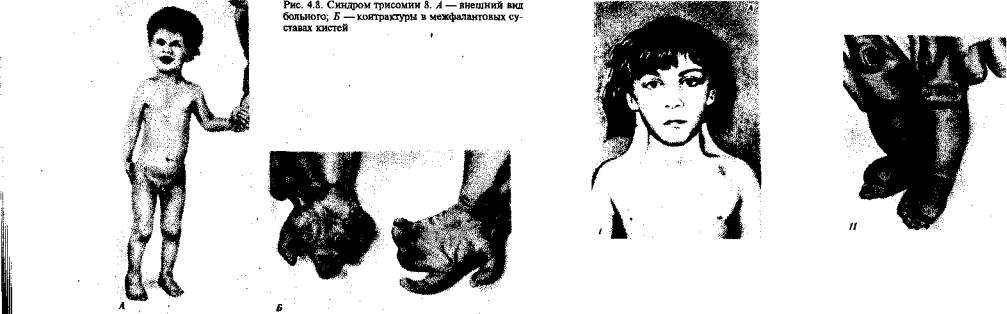
•І |
Среди других аутосомных трисомий известны трисомии по 13-й |
1| |
хромосоме — Синдром Патау (рис. 4.6), а также по 18-й хромосоме — |
•v синдром Эдвардса (рис. 4.7), при которых жизнеспособность новорож-
щденных резко снижена. Они гибнут в первые месяцы жизни из-за
Цмножественных пороков развития. Применение методов дифференци- ;
•ального окрашивания хромосом позволило открыть три новых синд-;
Црома, обусловленных трисомиями по 8, 9 и 22-й хромосомам, при І
•I |
которых также наблюдаются тяжелые комплексные пороки развития > |
I |
(рис. 4.8). |
• I |
Достаточно часто у человека встречаются анэуплоидии по половым; |
• I |
хромосомам (рис. 4.9—4.11). В отличие от анэуплоидии по аутосомам| |
Цг |
дефекты умственного развития у больных выражены не столь отчет-1 |
Иливо, у многих оно в пределах нормы, а иногда даже выше среднего. '•
•Вместе с тем у них постоянно наблюдаются нарушения развития
Вполовых органов и гормонозависимого роста тела. Реже встречаются]
•пороки развития других систем. Относительно благоприятные последа)
•ствия увеличения числа Х-хромосом, видимо, связаны с возможностью j
1 |
компенсации дозы соответствующих генов благодаря естественной!! |
|
1 |
генетической инактивации этих хромосом, а также мозаичному характеру} |
|
и |
такой инактивации. |
j |
Iі |
Среди анэуплоидных синдромов по половым хромосомам моносо- |
|
| |
мия X (ХО) {синдром Шерешевского — Тернера) встречается много реже, j |
|
1 |
186 |
|
I |
|
|
К h и |
п н і\ |
і |
|||
1 |
2 |
|
|
5 6 |
X |
II |
И II |
II |
It |
II |
|
|
|
|
10 |
|
12 |
ДІ |
| 4 |
А 6 |
XI |
| | |
| | |
13 |
14 |
15 |
16 |
17 |
18 |
* > |
«Я |
|
|
21 |
22 |
19 |
20 |
|
|
||
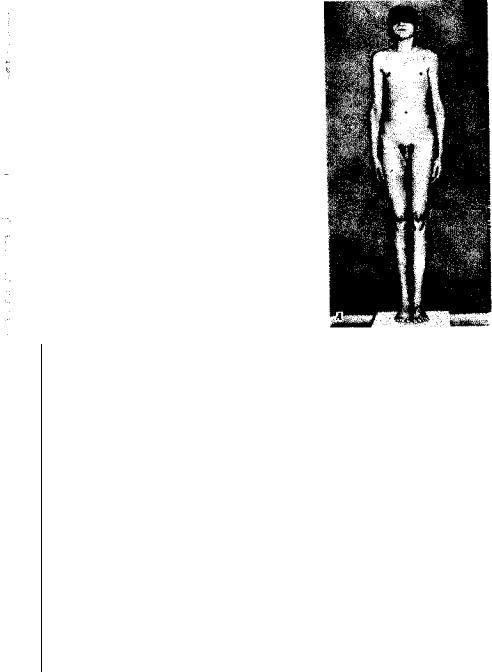
Рис. 4.9. Синдром моносомии X (ХО-синдром, синдром Шерешерского — Тернера). А —внешний вид больной; Б — кариотип женщины с синдромом ХО:
/ — выраженная трапециевидная шейная складка, широкая грудная клетка, широко расставленные, слаборазвитые соски молочных желез, II—характерные лимфатические отеки на ногах
ми I!
1 |
2 |
|
|
|
4 ' |
5 |
|
|
|
п а |
|
11 |
12 |
|
|
|
II |
П І» П |
М |
13 |
14 |
«ft |
|
|
|
|
|
||
*х де |
•я |
|
|
|
16 |
17 |
18 |
|
|
«ж |
ftt |
|
ш |
|
19 |
20 |
|
|
|
Aft |
А* |
|
|
|
21 |
22 |
|
|
|
РИС. 4.10. Кариотип женщины с синдромом трисомии X
чем трисомия X, синдром Клайнфельтера (XXY, XXXY), а также XYY, что указывает на наличие сильного отбора против гамет, не содержащихj половых хромосом, или против зигот ХО. Это предположение подтвер ждается достаточно часто наблюдаемой моносомией X среди спонтан- ; но абортированных зародышей. В связи с этим допускается, что выжившие зиготы ХО являются результатом не мейотического, a J митотического нерасхождения, или утраты Х-хромоеомы на ранних ! стадиях развития (см. рис. 4.9), Моносомии Y0 у человека не обнару жено.
Организмы с анэуплоидией по половым хромосомам при наличии | Y-хромосомы развиваются по мужскому типу и'фенотипически дают $ синдром Клайнфельтера (рис. 4.11). Это является еще одним свиде- •", тельством в пользу расположения фактора, определяющего мужской \ тип развития в Y-хромосоме. j
Из синдромов, связанных со структурными аномалиями хромосом, ' известен транслокационный синдром Дауна (см. рис. 4.5), при котором \ число хромосом в кариотипе формально не изменено и равно 46, так как дополнительная 21-я хромосома транслоцирована на одну из і акроцентрических хромосом. При транслокации длинного плеча 22-й
Рис. 4.11. Синдром Клайнфельте ра. А —^ внешний вид больного (характерен высокий рост, непро порционально длинные конечно сти); Б—кариотип больного (XXY)
Ufl ••• рп
! |
1 |
2 |
3 |
1 4 5 и nunпий
11 6 7 8 9 10 . 11 12
ДО ДА Aft
І |
13 |
14 |
15 |
| |
16 |
17 |
18 |
|
XX |
X» |
XXY |
|
19 |
20 |
|
і |
|
|
|
|
А А |
А* |
|
|
21 |
22 |
U |
188
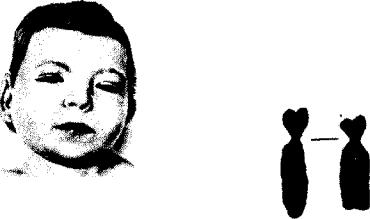
Рис. 4.12. Синдром хромосо мы 5р (синдром кошачьего крика). А — внешний вид больного; Б — деления корот кого плеча 5-й хромосы
хромосомы на 9-ю развивается хронический миелолейкоз. При делеции короткого плеча 5-й хромосомы развивается синдром кошачьего крика, при котором наблюдаются общее отставание в развитии, низкая масса при рождении, лунообразное лицо с широко расставленными глазами
ихарактерный плач ребенка, напоминающий кошачье мяукание, причиной которого является недоразвитие гортани (рис. 4.12).
Уносителей некоторых перицентрических инверсий нередко наблю даются аномалии в виде умственной отсталости той или иной степени
ипороков развития. Довольно часто такие перестройки наблюдаются
в9-й хромосоме человека, однако они существенно не влияют на развитие организма.
Таким образом, нарушение наследственной программы организма, развивающегося из аномальных гамет, или мозаицизм его клеток, связанный с соматическими мутациями, являются причиной либо гибели организма, либо более или менее выраженного снижения его ; жизнеспособности. !
РАЗДЕЛ III
ОНТОГЕНЕТИЧЕСКИЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИВОГО
В иерархической системе организации живого онтогенетический уровень тесно связан с другими уровнями. Элементарной единицей жизни здесь является особь, в процессе ее индивидуального развития. Реально существующие в природе организмы на протяжении жизни непосредственно взаимодействуют с окружающей средой —неживой природой, особями своего и других видов. В этом проявляется взаи мосвязь онтогенетического, популяционно-видового, биогеоценотического и биосферного уровней, в которые так или иначе включены отдельные организмы. В процессе взаимодействия особей с окружаю щей средой осуществляется отбор организмов, наиболее приспособ ленных в силу их наследуемых свойств. Основной задачей, решаемой на онтогенетическом уровне, является формирование организма, спо собного произвести потомство, передав ему наследственную програм му, на основе которой у нового поколения формируются характерные черты данного вида. При половом размножении эта задача решается не единичной особью, а в рамках популяции организмов данного вида, в которой находятся особи обоих полов.
Установив непосредственную связь онтогенетического уровня с надорганизменными уровнями организации жизни, нужно отметить, что для осуществления основной задачи — оставления потомства и обеспечения непрерывности существования вида — необходимо обес печить формирование зрелого в репродуктивном отношении орга низма и его жизнеспособность на всех стадиях онтогенеза. Это достигается благодаря функционированию элементарных единиц суборганизменных уровней организации—молекулярно-генетиче- ского и клеточного.
Изучение биологических закономерностей, действующих на онто генетическом уровне организации живого, представляет особый инте рес в системе медицинского образования, так как индивидуальное развитие человека в норме и при патологических отклонениях является важным объектом непосредственной врачебной деятельности.
191
ГЛАВА 5
РАЗМНОЖЕНИЕ
Среди многообразных проявлений жизнедеятельности (питание, обустройство местообитания, защита от врагов) размножению принад лежит особая роль. В известном смысле существование организма является подготовкой к выполнению им главной биологической задачи —участию в размножении. В основе способности организмов к размножению лежат определенные клеточные механизмы.
Продолжительность жизни особи короче продолжительности су ществования вида, к которому она принадлежит. Поэтому история вида — это история сменяющихся поколений организмов. Очередное (дочернее) поколение образуется в результате размножения особей предшествующего (родительского) поколения. Способность к размно жению является неотъемлемым свойством живых существ. С его помощью сохраняются во времени биологические виды и жизнь как таковая. Биологическая роль размножения состоит в том, что оно обеспечивает смену поколений. Различия, закономерно проявляющиеся в фенотипах особей разных поколений, делают возможным естествен ный отбор и, следовательно, эволюцию жизни.
Размножение возникло в ходе исторического развития органиче ского мира на самом раннем этапе вместе с клеткой. В процессе биологического размножения наряду со сменой поколений и Поддер жанием достаточного уровня внутривидовой изменчивости решаются также задачи увеличения числа особей, сохранения складывающихся в эволюции типов структурно-физиологической организации (путем восп роизведения себе подобного). Последнее связано с тем, что при размножении осуществляется передача в ряду поколений генетическо го материала (ДНК), т.е. определенной, специфичной для данного вида биологической информации.
5 . 1 . СПОСОБЫ И ФОРМЫ РАЗМНОЖЕНИЯ
Взависимости от характера клеточного материала, используемого
вцелях размножения, выделяют различные способы и формы по следнего. Различают два способа размножения: бесполое и половое (табл. 5.1).
Различные формы бесполого размножения представлены на сх. 5.1. Деление надвое приводит к возникновению из одного родительского организма двух дочерних. Оно является преобладающей формой у прокариот и простейших, но встречается и у многоклеточных: продоль ное у медуз, поперечное у кольчатых червей. Множественное деление (шизогония) встречается среди простейших, в том числе паразитов' человека (малярийный плазмодий). При размножении почкованием потомок формируется первоначально как вырост на теле родителя с
последующей его отшнуровкой (гидра). Фрагментация заключается в распаде тела многоклеточного организма на части, которые далее превращаются в самостоятельных№ особей (плоские черви, иглоко жие). У видов, размножающихся спорами, дочерний организм развииастся из специализированной клетки-споры.
Т а б л и ц а 5.1. Общая характеристика бесполого и полового размножения
Показатель |
|
|
|
|
Способ размножения |
|
|
|
|
|||
|
|
|
бесполое |
|
|
|
|
|
|
|
|
|
1. Клеточные |
источ |
Многоклеточные: одна или |
Родители |
образуют |
по |
|||||||
ники наследственной несколько соматических (те ловые |
клетки |
(гаметы), |
||||||||||
информации для разлесных) |
клеток |
родителя; специализированные |
к вы |
|||||||||
пития потомка |
|
одноклеточные: клетка-орга полнению |
функции |
раз |
||||||||
|
|
низм как целое |
|
|
множения. Родитель |
пред |
||||||
|
|
|
|
|
|
|
ставлен |
в потомке |
исходно |
|||
|
|
|
|
|
|
|
одной клеткой |
|
|
|
||
Родители |
|
Одна особь |
|
|
Обычно две особи |
|
|
|||||
Потомство |
|
Генетически точная копия |
Генетически отличны |
от |
||||||||
|
|
родителя, |
т.е. в отсутствие обоих родителей |
|
|
|
||||||
|
|
соматических мутаций |
клон |
|
|
|
|
|
|
|||
|
|
организмов |
|
|
|
|
|
|
|
|
||
Главный клеточный |
Митоз |
|
|
|
|
Мейоз |
|
|
|
|
||
механизм |
|
|
|
|
|
|
|
|
|
|
|
|
Эволюционное |
зна |
Способствует |
поддер |
За |
счет |
генетического |
||||||
чение |
|
жанию |
наибольшей |
при разнообразия создает |
пред |
|||||||
|
|
способленности в маломепосылки к освоению раз |
||||||||||
|
|
няющихся |
условиях |
оби нообразных |
условий |
обита |
||||||
|
|
тания, |
|
усиливает |
роль ния; дает эволюционные и |
|||||||
|
|
стабилизирующего |
естест экологические перспективы; |
|||||||||
|
|
венного отбора |
|
|
способствует |
осуществлению |
||||||
|
|
|
|
|
|
|
творческой |
роли |
естест |
|||
венного отбора
В зависимости от формы бесполого размножения потомок разви вается либо из одной клетки (спорообразование, шизогония, деление), либо из группы клеток родителя. В последнем случае размножение называют вегетативным. Оно распространено среди растений.
Бесполое размножение наблюдается у животных с относительно низким уровнем структурно-физиологической организации, к кото рым принадлежат многие паразиты человека. У паразитов бесполое размножение не только служит увеличению численности особей, но способствует расселению, помогает пережить неблагоприятные ус ловия.
5.2. ПОЛОВОЕ РАЗМНОЖЕНИЕ
Хотя в процессе развития жизни бесполое размножение возникло первым, половое размножение существует на Земле уже более 3 млрд. лет. Оно обнаруживается в жизненных циклах всех основных групп организмов. Распространенность полового размножения объясняется
192 |
13. Ярыгин. Т. 1. |
193 |
« о S
о
• О О о
litre Si
< 2 | § B £
|
I |
|
о |
£ |
8 , |
ex |
I
Is
Он
8
2 6 и
І s s
й g 2 »
§
і
о
§
тем, что оно обеспечивает значительное генетическое разнообразие и, следовательно, фенотипическую изменчивость потомства. Этим дости гаются большие эволюционные и экологические (расселение в разные среды) возможности.
В основе полового размножения лежит половой процесс, суть кото рого сводится к объединению в наследственном материале для развития потомка генетической информации от двух разных источников — родителей. Представление о половом процессе дает явление конъюга ции, например инфузорий. Он заключается во временном соединении двух особей с целью обмена (рекомбинации) наследственным матери алом. В результате появляются особи, генетически отличные от роди тельских организмов. В дальнейшем они осуществляют бесполое размножение. Поскольку количество инфузорий после конъюгации остается неизменным, говорить о размножении в прямом смысле нет оснований. У простейших половой процесс может осуществляться в виде копуляции, которая заключается в слиянии двух особей в одну, объединении и рекомбинации наследственного материала. Далее такая особь размножается делением. На определенном этапе эволюции у многоклеточных организмов половой процесс как способ обмена генетической информацией между особями в пределах вида оказался связанным с размножением.
Для участия в половом размножении в родительских организмах вырабатываются гаметы —клетки, специализированные к обеспече нию генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы —клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития.
У некоторых организмов зигота образуется в результате объедине ния гамет, не отличимых по строению. В таких случаях говорят об изогамии. У большинства видов по структурным и функциональным признакам половые клетки делятся на материнские (яйцеклетки) и
Рис. 5.1. Половой диморфизм на уровне половых клеток. А —яйцеклетка; Б —сперма тозоиды:
/ —цитоплазма, 2 — ядро, 3—хроматин ядра, 4—шейка, 5—жгутик, 6 — головка
13* |
195 |
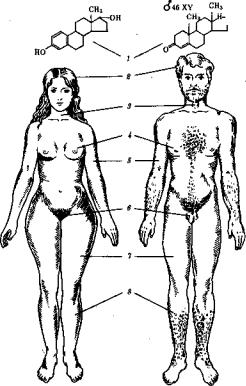
отцовские (сперматозоиды). Как правило, яйцеклетки и сперматозоиды ;| вырабатываются разными организмами —женскими (самки) и муж скими (самцы). В подразделении гамет на яйцеклетки и сперматозои ды, а особей на самок и самцов заключается явление полового, диморфизма (рис. 5.1; 5.2). Наличие его в природе отражает различия в задачах, решаемых в процессе полового размножения мужской или! женской гаметой, самцом или самкой. ?
Образование гамет обоих видов в одном организме, имеющем и!| мужскую, и женскую половые железы, называют гермафродитизмом1.
$ 4 6 XX
s-OH
Рис. 5.2. Половой диморфизм у людей на организменном уровне
Характерны различия по: /—кариотипу и главному половому гормону, 2—структуре волос и характеру оволосения, 3 —строению гортани, 4 —развитию молочных желез, 5 —развитию муску латуры, 6—строению половых органов, 7—распределению жировой ткани, 8 — показателям роста j| длинных трубчатых костей
От истинного гермафродитизма, о котором здесь идет речь, следует отличать гермафродитизм ложный, заключающийся в сочетании в одной особи наружных половых <! органов и вторичных половых признаков обоих полов при наличии половых желез одного типа — мужского или женского.
196
Гермафродитизм характерен для некоторых паразитов человека, на пример плоских червей. Несмотря на продукцию гермафродитами и мужских, и женских гамет, самооплодотворение для них нетипично, что связано обычно с несовпадением времени созревания яйцеклеток и сперматозоидов. Истинный гермафродитизм описан у человека. Чаще он развивается в результате нарушения эмбриогенеза при оди наковом наборе половых хромосом — XX или XY во всех соматических клетках. У некоторых людей-гермафродитов обнаружен мозаицизм по половым хромосомам. Одни соматические клетки имеют пару XX, другие — XY.
Хотя оплодотворение представляет собой характерный признак полового размножения, дочерний организм иногда развивается из неоплодотворенной яйцеклетки. Это явление называют девственным развитием или партеногенезом. Источником наследственного матери ала для развития потомка в этом случае обычно служит ДНК яйцек летки — гиногенез. Реже наблюдается андрогенез—развитие потомка из клетки с цитоплазмой ооцита и ядром сперматозоида. Ядро женской
гаметы в случае андрогенеза погибает.
Обязательный партеногенез является измененной формой полового размножения в эволюции некоторых видов животных. У пчел, напри мер, он используется как механизм генотипического определения пола: женские особи (рабочие пчелы и царицы) развиваются из оплодотворен ных яйцеклеток, а мужские (трутни) — партеногенетически. Партеноге нез включен в жизненные циклы многих паразитов. Он обеспечивает рост численности особей в условиях, затрудняющих встречу партнеров противоположного пола. Имеются указания на возможность девствен ного развития у человека. В яичниках девушек, погибших при случай ных обстоятельствах, в отсутствие предшествующего осеменения нахо дили зародыши на ранних этапах дробления. Невозможность завершен ного партеногенеза у людей в настоящее время доказана и связана с не обходимостью наличия обоих геномов, мужского и женского (см. с. 250—-251). Наблюдения завершенного эмбриогенеза с партеногенетическим развитием в отношении человека отсутствуют. При партеногенезе, как и при типичном половом размножении, развиваются особи с дипло идными соматическими клетками. Восстановление диплоидного набора хромосом происходит обычно путем слияния ооцита и редукционного тельца во втором делении мейоза.
У некоторых видов закономерно наблюдается полиэмбриония — бесполое размножение зародыша, возникающего путем полового раз множения. Полиэмбриония, к примеру, типична для броненосцев и заключается в разделении на стадии бластулы клеточного материала первоначально одного зародыша между 4—8 зародышами, из которых развиваются полноценные особи. В результате полиэмбрионии у че ловека рождаются однояйцевые близнецы.
197
5.2.1.Чередование поколений с бесполым
иполовым размножением
Большинство организмов, обычно размножающихся бесполым пу- j тем, способно к половому размножению. При этом ряд поколений с бесполым размножением сменяется поколением особей, размножаю-І щихся с помощью гамет или же осуществляющих половой процесс] Вслед за этим вновь наблюдается бесполое размножение. Смена (череА дование) половых и бесполых поколений происходит у разных видов с разной периодичностью, регулярно или через неодинаковые промер жутки времени. j
Первичное чередование поколений заключается в смене полового;) размножения спорообразованием. Оно наблюдается у представителей;] классов споровиков, жгутиконосцев, некоторых растений и отражает! сохранение в филогенезе соответствующих организмов как болеедревней (бесполой), так и более прогрессивной (половой) форм раз-; множення. Вторичное чередование поколений заключается в переходе на|| некоторых стадиях жизненного цикла к бесполому или партеногенез тическому размножению животных, освоивших половое размножение. | Оно распространено у кишечнополостных, членистоногих. !|
Включение в цикл развития организмов, размножающихся преиму- j щественно бесполым путем, полового поколения время от времени | активизирует комбинативную изменчивость и этим способствует пре- :| одолению генетического однообразия потомков, расширяя эволюци-; онные и экологические перспективы группы.
5.3. ПОЛОВЫЕ КЛЕТКИ
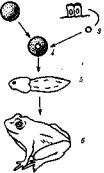
"По сравнению с другими клетками функция гамет уникальна. Они • обеспечивают передачу наследственной информации между особями , разных поколений, чем сохраняют жизнь во времени. Был период в ' биологии, когда половые и соматические клетки противопоставляли { друг другу, наделяя лишь первые всей полнотой свойств жизни, проносимых ими через поколения. В настоящее время эксперимен тально доказана возможность развития полноценного организма на [ основе наследственной информации ядра дифференцированной сома-: тической клетки, например кишечного эпителия (рис. 5.3).
В 1990-х годах группе английских ученых, использовавших принципиально анало- ; гичныи подход, удалось вырастить (клонировать) овцу Долли. Это был пример успешного [ решения задачи получения взрослой особи на основе генетического материала сомати-'! ческой клетки у млекопитающих. Напомним, что клоны представлены генетически идентичными клетками, образовавшимися путем последовательных мйтотических деле ний исходно одной клетки-родоначальницы и ее потомков. Вытекающая из этого техническая возможность клонирования людей порождает серьезные морально-этиче ские проблемы, так как допускает преднамеренное «тиражирование» избранного чело века с присущим только ему набором физических психо-эмоциональных свойств и даже интеллектуального потенциала. В настоящее время общественностью, в том числе и научной, признается недопустимым клонирование людей. Вместе с тем, использование
198
Рис. 5.3. Опыт, показывающий функциональную полно |
1 |
2 |
ценность наследственного материала дифференцирован ной клетки:
/ —яйцеклетка с убитым УФ-лучами ядром —источник цитоп лазмы, 2 —эпителиальные клетки кишечника головастика —ис точник наследственного материала, 3 —ядро, 4 —пересадка ядра из эпителиальной клетки в яйцеклетку, 5 — головастик, 6 — ля гушка
технологии клонирования для получения достаточного числа клеток определенного типа, генетически неотличимых от клеток данного организма и, следовательно, иммунологически с ним совместимых, рассматривается как перспективное направление в области трансплантации тканей.
Гаметы представляют собой одно из многих направлений дифференцировки клеток много
клеточного организма. Они образуют «клеточную линию», специали зированную к выполнению репродуктивной функции. Предполагают, что клетки указанной линии возникают из бластомеров, имеющих на вегетативном полюсе цитоплазму особого рода—так называемую
зародышевую плазму1,богатую РНК.
В сравнении с другими линиями соматических клеток (эпители альные, нервные, мышечные) гаметы характеризуются рядом отличий. Важнейшее из них — гаплоидный набор хромосом в ядрах, что обеспе чивает воспроизведение в зиготе типичного для организмов данного вида диплоидного числа хромосом.
Действительно, оплодотворение сперматозоидом яйцеклетки, ядра которых содержат по 23 хромосомы, обусловливает формирование зиготы с 46 хромосомами, что типично для соматических клеток человека. Гаметы отличаются необычным для других клеток значением
ядерно-цитоплазматического отношения. У яйцеклеток оно снижено благодаря увеличенному объему цитоплазмы, в которой размещен питательный материал (желток) для развития зародыша. У спермато зоидов благодаря малому количеству цитоплазмы ядерно-цитоплазма- тическое отношение высокое. Это находится в соответствии с главной функциональной задачей мужской гаметы — транспортировкой на следственного материала к яйцеклетке.
Половые клетки отличаются низким уровнем обменных процессов,
близким к состоянию анабиоза. Мужские гаметы не вступают в митотический цикл. У яйцеклеток эта способность восстанавливается при оплодотворении или действии фактора, активирующего партено генез.
По ряду признаков женские и мужские гаметы отличаются друг от друга, что связано с различными функциями яйцеклетки и спермато-
Термин зародышевая плазма был ранее использован А. Вейсманом, но в совершенно ином смысле — для обозначения наследственного вещества ядра клетки (хромосом).
199