

Тема 6 Молекулярные основы проведения нервного импульса в нервных волокнах и синапсах
Природа создала два принципиально различных способа межклеточной сигнализации. Один из них состоит в том, что сообщения передаются при помощи электрического тока; во втором используются молекулы, передаваемые от одной клетки к другой (гормоны). В обоих случаях передача сигнала зависит от проницаемости мембран.
Электрическая система передачи информации служит для передачи нервного раздражения и осуществляется специальными нервными клет ками – нейронами. Существуют нейроны двух видов: чувствительные (передают высшим центрам нервной системы импульсы, возникающие на рецепторных мембранах под влиянием внешних раздражителей) и двигательные (передают импульсы от высших центров нервной системы к мышечным клеткам и секреторным органам).
Любая нервная клетка состоит из тела и отростков различной длины. Тело клетки содержит все органеллы, присущие эукариотическим клеткам (ядро, митохондрии, ЭПР и др.), и служит биосинтетическим центром. Дендриты представляют собой систему ветвящихся отростков различной длины, которые отходят от тела нейрона и увеличивают поверхность, способную принимать сигналы от других клеток (рис.6.1).
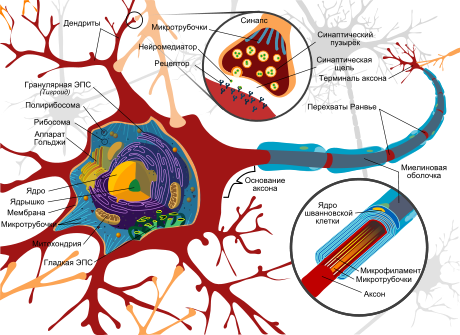
Рис.6.1 Строение нервной клетки
Самый длинный из отростков называется аксоном; в процессе развития нервной клетки один из ее отростков начинает расти до тех пор, пока не достигает той точки, с которой ему предстоит поддерживать контакт. Длина аксона достигает размеров от долей миллиметра до 1.5 м (у человека). Аксон подобен телеграфному кабелю тем, что он хорошо изолирован. Изоляцию вокруг аксона создают шванновские клетки. Плазматическая мембрана этих клеток концентрическими слоями (до 100 слоев) плотно наматывается на аксон, образуя сегмент миелиновой оболочки длиной около 1 мм. Миелиновая оболочка состоит из липидно-белковых мембран, плохо пропускающих ионы, что практически полностью предотвращает утечку тока из аксона. Между смежными сегментами миелиновой оболочки остаются небольшие открытые участки аксона длиной около 1 мкм – перехваты Ранвье. Сигналы, проводимые нейронами, передаются от одной клетки к другой в местах контакта, называемых синапсами (места плотного контакта) (рис.5.7). Изменение электрического потенциала в пресинаптической мембране приводит к высвобождению нейромедиатора, который выходит через синаптическую щель и связывается с рецептором на постсинаптической мембране, что приводит к изменению электрофизиологического состояния постсинаптической клетки. Происходит превращение электрического сигнала в химический и далее химического – вновь в электрический.
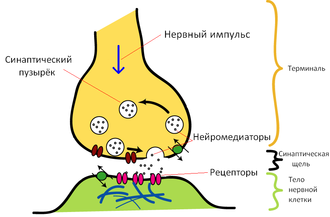
Рис.6.2 Синапс
Причина возникновения электрического сигнала состоит в изменении электрического потенциала на плазматической мембране нейрона. Передача сигналов основана на том, электрическое возмущение, возникшее в одном участке клетки, распространяется на другие участки. Если нет дополнительного усиления, эти возмущения затухают по мере удаления от их источников. На коротких расстояниях затухание незначительно, и многие нейроны проводят сигналы пассивно, без усиления.
Однако, для дальней связи такого пассивного распространения сигнала недостаточно, и поэтому у нейронов с длинными отростками в ходе эволюции выработался активный сигнальный механизм. Электрический стимул, сила которого превышает определенную пороговую величину, вызывает взрыв электрической активности, распространяющийся с большой скоростью вдоль плазматической мембраны нейрона. Эту бегущую волну возбуждения называют потенциалом действия или нервным импульсом. Потенциал действия передает информацию с одного конца нейрона на другой без затухания со скоростью до 100 м/с, а в некоторых случаях еще быстрее.
Механизм передачи нервного импульса.
1. В состоянии покоя (при отсутствии возбуждения) между внутрен-
ней и наружной сторонами плазматической мембраны аксона поддерживается разность потенциалов (трансмембранный потенциал ∆Ψ), называемая потенциалом покоя, при котором суммарный ток различных ионов, пересекающих мембрану, равен нулю. Внутри мембраны присутствует отрицательный электрический заряд, ∆Ψ = – 70 мВ. При повышении потенциала до значения –60 мВ происходит возбуждение аксона, открываются потенциалзависимые каналы для катионов Na+ и K+ (Na+-канал считается быстрым, а K+-канал медленным).
2. В начальный момент в мембрану аксона проникают в основном ионы Na+, что приводит к резкому увеличению мембранного потенциала и достижению его максимального значения. Теперь для дальнейшего проникновения внутрь аксона ионам Na+ пришлось бы преодолевать градиент потенциала, поэтому поступление Na+ из наружной среды прекращается и в этот момент Na+-канал как бы закрывается.
3. Через K+-канал во внешнюю среду вытекают ионы K+ по своему концентрационному градиенту до тех пор, пока внутри аксона не восстанавливается первоначальный отрицательный потенциал. Однако, процесс поступления K+ во внешнюю среду продолжается несколько дольше, чем требовалось бы, и поэтому потенциал падает до уровня меньшего, чем значение потенциала покоя. В этот момент аксон становится невозбудимым, т.е. лишается способности проводить нервный импульс.
4. После этих событий первоначальное значение потенциала очень быстро (тысячные доли секунды) восстанавливается, и аксон вновь готов к дальнейшему проведению импульса.
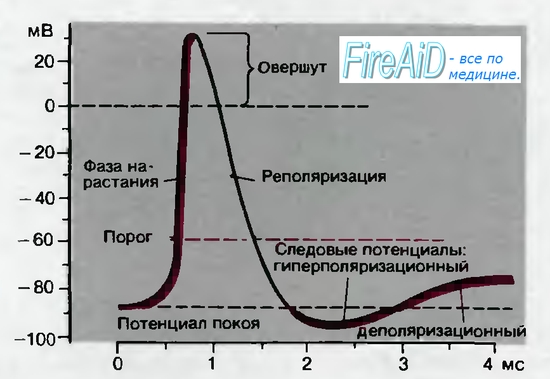
Рис.6.3. Потенциал действия
Аксон изолирован слоем шванновских клеток, подобных изоляционной ленте вокруг проводника, эта миелиновая оболочка не пропускает катионы. В результате перехода внутрь аксона большого количества ионов Na+, в аксоне возникают продольные токи, т.е. ионы Na+ мигрируют в следующий соседний отсек аксона, находящийся в состоянии покоя.
Потенциал в этом отсеке повышается, при достижении его значения – 60 мВ открываются Na+-каналы, потенциал достигает максимального значения и повторяется вся приведенная выше цепь событий в аксоне.
Таким образом, по всей длине аксона распространяется волна потенциала действия. Этот процесс аналогичен тому, как огонь бежит по бикфордову шнуру. Скорость распространения - от 1 до 100 м/с в зависимости от типа аксона.
Потенциал действия достигает конца аксона и дает сигнал для выброса в синаптическую щель нейромедиатора, который в следующей клетке возбуждает ту же цепь событий. После проведения импульса система возвращается в первоначальное равновесное состояние, градиенты концентраций катионов Na+ и K+ восстанавливает фермент Na+,K+-АТФаза.
Таким образом, за проведение нервного импульса отвечает изменение проницаемости мембраны аксона. Экспериментальные данные подтвердили тот факт, что катионы Na+ и K+ проходят через мембрану нерва по разным каналам. Так, действие яда тетродотоксина, парализующего проведение нервного импульса, блокирует проводимость Na+ -каналов, в то время как проводимость ионов K+ не затрагивается. Было подсчитано число Na+-каналов в аксонах кролика – оно не превышает 75 каналов на 1 мкм2, скорость движения Na+ соответствует механизму простой диффузии через белковый канал (108 с-1). Установлено, что число K+-каналов много больше, но меньше проводимость этих ионов, что соответствует механизму транспорта ионов K+ с помощью специальных переносчиков.
Специальные механизмы транспорта веществ через биомембрану (эндо- и экзоцитоз)
Транспортные белки обеспечивают проникновение через клеточные мембраны многих полярных молекул небольшого размера, однако они не способны транспортировать макромолекулы, например, белки, полинуклеотиды или полисахариды. Тем не менее, в большинстве клеток определенные макромолекулы могут проходить в двух направлениях через плазматические мембраны, а некоторые клетки даже способны поглощать большие клеточные частицы. Механизмы этих процессов значительно отличаются от механизмов транспорта небольших молекул или ионов.
Экзоцитоз представляет собой механизм секреции макромолекул из клетки во внешнюю среду.
При переносе макромолекул во внешнюю среду происходит последовательное образование и слияние окруженных мембраной пузырьков (везикул) с плазматической мембраной (рис.6.3):

Рис. 6.3 Схема экзоцитоза
Например, для секреции инсулина клетки упаковывают его во внутриклеточные пузырьки, которые сливаются с плазматической мембраной и открываются во внешнеклеточное пространство, высвобождая при этом молекулы инсулина. Во всех эукариотических клетках имеются специальные везикулы, переносящие аналогичным образом от аппарата Гольджи к плазматической мембране ее вновь синтезированные компоненты. Таким образом, путь синтезированных в клетке молекул – это адсорбция на поверхности клетки в качестве ее новых компонентов, либо экзоцитоз в интерстициальную жидкость или в кровь питательных веществ и сигнальных молекул.
Процесс слияния двух мембран состоит из двух фаз и начинается с того, что эти мембраны приходят в близкое соприкосновение друг с другом (слипание бислоев). Электронная микрофотография регистрирует при этом появление пятислойной мембранной структуры, однако ее существование кратковременно. Затем слипшиеся бислои быстро перестраиваются и объединяются, образуя одну непрерывную мембрану, вследствие чего секреторные пузырьки открываются во внеклеточное пространство. Процессы слипания и объединения бислоев представляют собой фундаментальные процессы, лежащие в основе эндоцитоза и экзоцитоза, деления и слияния клеток.
Индуцированный экзоцитоз.
Было установлено, что одни вещества непрерывно секретируются клетками, тогда как другие запасаются в секреторных пузырьках и высвобождаются только после получения клеткой соответствующего сигнала извне. Этот сигнал к секреции часто представляет собой химический медиатор, например, гормон, связывающийся с рецепторами на поверхности клетки. В результате происходит активация рецепторов, которая вызывает обычно кратковременное повышение концентрации свободных ионов Cа2+ в цитозоле, а это в свою очередь индуцирует процесс экзоцитоза. Представляется вероятным, что ионы Cа2+ действуют в ограниченной области секретирующей клетки, инициируя локальный ответ части цитоплазмы и располагающейся над ней плазматической мембраны. Этот вывод был сделан в опытах с тучными клетками, секретирующими гистамин в ответ на связывание специальных лигандов с рецепторами на их поверхности. Если тучные клетки инкубировали в среде, содержащей растворенный стимулятор, то экзоцитоз наблюдался по всей клеточной поверхности. В другом случае стимулятор был закреплен на подложке, и экзоцитоз ограничивался местом контакта с подложкой.
Эндоцитоз – клетки способны также поглощать макромолекулы и частицы, используя в обратной последовательности сходный механизм:
поглощенное вещество постепенно окружается небольшим участком плазматической мембраны, которая сначала впячивается, а затем отщепляется, образуя внутриклеточный пузырек, содержащий захваченный клеткой материал.
В зависимости от размера образующихся пузырьков различают 2 типа процесса эндоцитоза:
1) пиноцитоз – поглощение жидкости и (или) растворенных веществ с помощью небольших пузырьков;
2) фагоцитоз – поглощение больших частиц, таких как микроорганизмы или обломки клеток. Большинство эндоцитозных пузырьков в конце концов сливается с первичными лизосомами. При этом образуются вторичные лизосомы, в которых переваривается большая часть макромолекулярного содержимого пузырьков. После этого основная часть мембранных компонентов пузырьков каким-то образом возвращается в плазматическую мембрану.
Эндоцитоз, также как и экзоцитоз, представляет собой локальную ответную реакцию плазматической мембраны. Большинство животных клеток непрерывно осуществляют эндоцитоз фрагментов своей плазматической мембраны. Таким путем поглощается также внеклеточная жидкость и растворенные в ней вещества.
При эндоцитозе одни пузырьки образуются из гладких участков плазматической мембраны, а другие – из ее окаймленных участков, называемых окаймленными ямками. Согласно данным электронной микроскопии эти участки со стороны цитоплазмы окружены щетинообразной каймой. Окаймленные ямки и пузырьки обеспечивают протекание процесса, называемого опосредуемый рецепторами эндоцитоз или адсорбционный эндоцитоз: поглощаемые молекулы связываются со специфическими белками-рецепторами, локализованными в окаймленных ямках (рис.5.7).
Этот процесс представляет собой избирательный концентрирующий механизм, позволяющий клеткам захватывать большие количества специфических лигандов без поглощения соответственно большого объема внеклеточной жидкости.
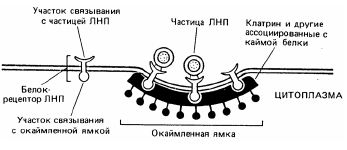
Рис.6.4 Схема адсорбционного эндоцитоза холестерина
Примером данного процесса служит поглощение животными клетками холестерина из внеклеточной среды, за счет этого обеспечивается большая часть потребности клеток в холестерине для синтеза мембран. Если этот процесс по какой-то причине заблокирован, то холестерин накапливается в крови и увеличивает риск атеросклероза. Основная часть холестерина переносится кровью в виде комплексов с белком, которые известны под названием - ЛНП - липопротеины низкой плотности. Эти комплексы представляют собой большие сферические частицы (22 нм в диаметре), каждая из которых имеет сердцевину, заполненную 1500 молекулами холестерина, связанного сложноэфирными связями с длинными цепями жирных кислот. Сердцевина ЛНП окружена липидным бислоем, содержащим белок одного-единственного типа.
Фагоцитоз у простейших организмов представляет собой форму питания, у млекопитающих большинство клеток не способно эффективно поглощать крупные частицы. Эту роль у млекопитающих выполняют 2 класса лейкоцитов – макрофаги и полиморфноядерные лейкоциты, защищая организм от вторгшихся микроорганизмов.
Важная особенность как экзоцитоза, так и эндоцитоза состоит в том, что секретируемые или поглощаемые макромолекулы локализуются в мембранных пузырьках и не смешиваются с другими макромолекулами или органеллами клетки. С помощью неизвестного механизма каждый пузырек сливается только со специфическими мембранными структурами, что гарантирует правильный перенос макромолекул.
Экзоцитоз и эндоцитоз не повторяют друг друга в обратном порядке из-за наличия стадии слипания бислоев, и благодаря такому различию, по-видимому, могут регулироваться независимо друг от друга.