

Ответы на вопросы:
Краткая форма (Нейрон, его физиологические свойства):
Нейрон — это нервная клетка с отростками, являющаяся основной структурной и функциональной единицей нервной системы. Она имеет строение, сходное с другими клетками: оболочка, протоплазма, ядро, митохондрии, рибосомы и другие органоиды. В нейроне различают три части: тело клетки — сома, длинный отросток — зксон и множество коротких разветвленных отростков — дендритов. Сома выполняет обменные функции, дендриты специализируются на приеме сигналов из внешней среды или от других нервных клеток, аксон на проведении и передаче возбуждения в область, удаленную от зоны дендритов. Аксон оканчивается группой концевых разветвлений для передачи сигналов другим нейронам или органам-исполнителям. Наряду с общим сходством в строении нейронов наблюдается большое разнообразие, обусловленное их функциональными различиями
Полная форма:
Нейрон – анатомо-гистологическая единица ЦНС. Он состоит из тела и отростков. Тела нейронов составляют серое вещество головного мозга. Их функции заключаются в переработке и хранении информации, а также в питании отростков.
Отростки нейронов:
аксоны – длинные маловетвистые отростки, проводящие информацию от тела нейрона к периферии
дендриты – короткие, сильноветвистые отростки, передающие информацию от периферии к телу нейрона.
Функции отростков заключаются в проведении информации к телу и от тела нейрона, в обеспечении взаимодействия нейронов с другими структурами.
По локализации нейроны подразделяются на центральные и периферические. Центральными называются те нейроны, тела которых лежат в пределах ЦНС. Периферические нейроны принадлежат периферической нервной системе. Они могут залегать в спинно-мозговых ганглиях, в ганглиях черепно-мозговых нервов, в ганглиях вегетативной нервной системы.
В зависимости от выполняемой функции нейроны делятся на 3 основные группы:
афферентные (чувствительные)
эфферентные (двигательные)
вставочные (контактные).
Афферентные нейроны обеспечивают восприятие раздражения и передачу информации в ЦНС.
Эфферентные нейроны обеспечивают передачу информации от ЦНС на периферию.
Вставочные нейроны обеспечивают передачу информации внутри ЦНС (с афферентных нейронов на эфферентные).
В зависимости от эффекта вставочные нейроны подразделяются на:
возбуждающие – оказывают возбуждающее влияние на эфферентные нейроны
тормозные – оказывают тормозное влияние на эфферентные нейроны.
В зависимости от вида медиатора в синапсе нейрона различают:
холинергические нейроны (медиатор – ацетилхолин)
адренергические нейроны (медиаторы – адреналин и норадреналин)
Среди множества известных функций нейрона: трофической, генераторной, проводящей возбуждение, наибольший интерес представляет способность нейронов синтезировать и секретировать биологическиактивные вещества. Одна нервная клетка может синтезировать только один вид медиатора.
Аксонный транспорт заключается в передвижении от тела клетки к окончанию аксона синтезированных в клеточных органеллах секреторных гранул. Помимо прямого аксонного транспорта, существует и обратный (ретроградный), сущность которого заключается в захвате аксонными окончаниями (путем пиноцитоза) из синаптической щели неизрасходованных молекул вещества и сохранении их в теле нейрона в виде некоторого резерва.
Понятие о нейроглии.
Нейроглия– это клетки, окружающие нейроны и входящие вместе с ними в состав ЦНС и ПНС. Количество глиальных клеток на порядок выше количества нервных клеток.
Функции нейроглии:
опорная – поддерживает нервные клетки
изолирующая – препятствует переходу нервных импульсов с тела одного нейрона на тело другого
регуляторная – участвует в регуляции работы ЦНС, в частности, обеспечивая передачу импульсов в нужном направлении
трофическая – участвует в обменных процессах нейронов
регуляторная – регулирует возбудимость нервных клеток.
(Механизм возникновения потенциала покоя)
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил:
1) силы диффузии;
2) силы электростатического взаимодействия. Значение электрохимического равновесия:
1) поддержание ионной асимметрии;
2) поддержание величины мембранного потенциала на постоянном уровне.
В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентра-ционно-электрохимическим.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ.
(Механизм возникновения потенциала покоя)
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания.
Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Лавинообразное поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит.
В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный.
(Изменение возбудимости при похождении волны возбуждения)
При развитии потенциала действия происходит изменение возбудимости ткани, причем, это изменение протекает но фазам (рис. 2). Состоянию исходной поляризации мембраны, которую отражает мембранный потенциал покоя, соответствует исходное состояние ее возбудимости и, следовательно, клетки - это нормальный уровень возбудимости. В период предспайка возбудимость ткани повышена, эта фаза возбудимости получила название первичной экзальтации. Во время развития предспайка мембранный потенциал покоя приближается к критическому уровню деполяризации и для достижения последнего достаточна сила раздражителя меньшая, чем пороговая (подпорого-вая).
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Она длится до конца перезарядки мембраны. Абсолютная рефрактерность, т. е. полная невозбудимость мембраны возникает в связи с тем, что натриевые каналы в начале полностью открываются, а затем инактивируются.
После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня - фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых.
Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости - фаза вторичной экзальтации. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, но сравнению с состоянием покои (исходной поляризацией) , то порог раздражения снижен, т. е. возбудимость повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы неполностью. В период развития положительного следового потенциала возбудимость ткани понижена - фаза вторичной рефрактерности. В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны) , удаляясь от критического уровня деполяризации, порог раздражения повышается и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Гиперполяризация мембраны развивается вследствие трех причин: во-первых, продолжающимися выходом ионов калия; во-вторых, открытием, возможно, каналов для хлора и поступление этих ионов в цитоплазму клетки; в-третьих, усиленной работой натрий-калиевого насоса.
(Проведение возбуждения по нервным волокнам)
Механизм распространения возбуждения у различных нервных волокон неодинаков. По современным представлениям распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия.
При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи,, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденным, заряженным положительно, вызывают деполяризацию мембраны до критического уровня с последующей генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот процесс повторяется многократно. На всем протяжении нервного волокна происходит процесс репродукции нового ПД в каждой точке мембраны волокна. Такое проведение возбуждения называете и непрерывным.
Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье) создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Местные электрические токи возникают между соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится заряженной отрицательно по отношению к поверхности соседнего невозбужденного перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным или скачкообразным. Скорость такого способа проведения возбуждения значительно выше и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
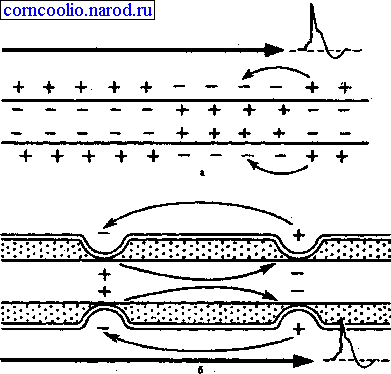
Рис. 4. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
"Перепрыгивание" потенциала действия через участок между перехватами возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не только через один, но и через два межперехватных промежутка. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др.
Нервные волокна обладают лабильностью - способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной ре-фрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную.
В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100) , то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции.
В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютнойрефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности.
Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инак-тивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется.
(Синапсы, их виды, структурные особенности)
Физиология синапсов.
В ЦНС нервные клетки связаны друг с другом посредством синапсов. Синапс – это структурно функциональное образование, которое обеспечивает передачу возбуждения или торможения с нервного волокна на иннервируемую клетку.
Синапсы по локализации делятся на центральные (расположены в пределах ЦНС, а также в ганглиях вегетативной нервной системы) и периферические (расположены вне ЦНС, обеспечивают связь с клетками иннервируемой ткани).
В функциональном отношении синапсы делятся на возбуждающие, в которых в результате деполяризации постсинаптической мембраны генерируется возбуждающий постсинаптический потенциал, и тормозные, в пресинаптических окончаниях которых выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала.
По механизму передачи синапсы делятся на химические и электрические. Химические синапсы передают возбуждение или торможение за счет особых веществ – медиаторов. В зависимости от вида медиатора химические синапсы подразделяются на:
холинергические (медиатор – ацетилхолин)
адренергические (медиаторы – адреналин, норадреналин)
По анатомической классификации синапсы делятся на нейросекреторные, нервно-мышечные и межнейронные.
Синапс состоит из трех основных компонентов:
пресинаптической мембраны
постсинаптической мембраны
синаптической щели
Пресинаптическая мембрана является окончанием отростка нервной клетки. Внутри отростка в непосредственной близости от мембраны имеется скопление пузырьков (гранул), содержащих тот или иной медиатор. Пузырьки находятся в постоянном движении.
Постсинаптическая мембрана является частью клеточной мембраны иннервируемой ткани. Постсинаптическая мембрана в отличие от пресинаптической имеет белковые хеморецепторы к биологически активным (медиаторам, гормонам), лекарственным и токсическим веществам. Важная особенность рецепторов постсинаптической мембраны – их химическая специфичность, т.е. способность вступать в биохимическое взаимодействие только с определенным видом медиатора.
Синаптическая щель представляет собой пространство между пре- и постсинаптичекой мембранами, заполненное жидкостью, близкой по составу к плазме крови. Через нее медиатор медленно диффундирует от пресинаптической мембраны к постсинаптической.
Особенности строения нервно-мышечного синапса обусловливают его физиологические свойства.
Одностороннее проведение возбуждения (от пре- к постсинаптической мембране), обусловленное наличием чувствительных к медиатору рецепторов только в постсинаптической мембране.
Синаптическая задержка проведения возбуждения (время между приходом импульса в пресинаптическое окончание и началом постсинаптического ответа), связанная с малой скоростью диффузии медиатора в синаптическую щель по сравнению со скоростью прохождения импульса по нервному волокну.
Низкая лабильность и высокая утомляемость синапса, обусловленная временем распространения предыдущего импульса и наличием у него периода абсолютнойрефрактерности.
Высокая избирательная чувствительность синапса к химическим веществам, обусловленная специфичностью хеморецепторов постсинаптической мембраны.
Этапы синаптической передачи.
Синтез медиатора. В цитоплазме нейронов и нервных окончаний синтезируются химические медиаторы – биологически активные вещества. Они синтезируются постоянно и депонируются в синаптических пузырьках нервных окончаний.
Секреция медиатора. Высвобождение медиатора из синаптических пузырьков имеет квантовый характер. В состоянии покоя оно незначительно, а под влиянием нервного импульса резко усиливается.
Взаимодействие медиатора с рецепторами постсинаптической мембраны. Это взаимодействие заключается в избирательном изменении проницаемости ионоселективных каналов эффекторной клетки в области активных центров связывания с медиатором. Взаимодействие медиатора со своими рецепторами может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. В случае увеличения проницаемости натриевых и кальциевых каналов усиливается поступление Na и Ca в клетку с последующей деполяризацией мембраны, возникновением ПД и дальнейшей передачей нервного импульса. Такие синапсы называются возбуждающими. Если повышается проницаемость калиевых каналов и каналов для хлора, наблюдается избыточный выход К из клетки с одновременной диффузией в нее Cl , что приводит к гиперполяризации мембраны, снижению ее возбудимости и развитию тормозных постсинаптических потенциалов. Передача нервных импульсов затрудняется или совсем прекращается. Такие синапсы называются тормозными.
Рецепторы, взаимодействующие с АХ, называются холинорецепторами. В функциональном отношении они разделяются на две группы: М - и Н-холинорецепторы. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, тогда как в мышцах внутренних органов – преимущественно М-холинорецепторы.
Рецепторы, взаимодействующие с НА, называются адренорецепторами. В функциональном отношении они делятся на альфа- и бета-адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие НА является деполяризующим, если он взаимодействует с альфа-адренорецепторами (сокращение мышечной оболочки стенок кровеносных сосудов или кишечника), или тормозным – при взаимодействии с бета-адренорецепторами (их расслабление).
Инактивация медиатора. Инактивация (полная потеря активности) медиатора необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня МП. Наиболее важным путем инактивации является гидролитическое расщепление с помощью ингибиторов. Для АХ ингибитором является холинэстераза, для НА и адреналина – моноаминоксидаза и катехолоксиметилтрансфераза.
Другой путь удаления медиатора из синаптической щели – «обратный захват» его пресинаптическими окончаниями (пиноцитоз) и обратный аксонный транспорт, особенно выраженный для катехоламинов.
В основе координационной деятельности ЦНС лежит взаимодействие процессов возбуждения и торможения.
Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани.
Торможение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся снижением функций ткани.
Первичное торможение в ЦНС возникает за счет тормозных нейронов. Это особый вид вставочных нейронов, которые при передаче импульса выделяют тормозной медиатор. Различают 2 вида первичного торможения: постсинаптическое и пресинаптическое.
Постсинаптическое торможение возникает, если аксон тормозного нейрона образует синапс с телом нейрона и, выделяя медиатор, вызывает гиперполяризацию клеточной мембраны, тормозя активность клетки.
Пресинаптическое торможение возникает, когда аксон тормозного нейрона образует синапс с аксоном возбуждающего нейрона, препятствуя проведению импульса.
(Спинной мозг, его функции, участие в регуляции мышечного тонуса)
Спинной мозг выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями.
Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, т.е. афферентными, передние двигательными, эфферентными. Эта закономерность называется законом Белла-Мажанди. Корешки каждого сегмента иннервируют 3 метамера тела, но в результате перекрывания каждый метамер иннервируется тремя сегментами. Поэтому при поражении передних корешков одного сегмента, двигательная активность соответствующего метамера лишь ослабляется.
Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все его нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов вегетативной нервной системы. Мотонейроны, в зависимости от функционального значения, разделяются на альфа – и гамма-мотонейроны. К a-мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных, т.е. рецепторных мышечных клеток. Тела a-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает.
Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС.
Нейроны симпатического отдела вегетативной нервной системы находятся в боковых рогах грудных сегментов, а парасимпатического в крестцовом отделе.
Проводниковая функция состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие. Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища.
К восходящим путям относятся несколько трактов. Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности.
Нисходящие пути также образованы несколькими трактами. Кортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры к a-мотонейронам спинного мозга. За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга к a-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к a-мотонейронам мышц разгибателей. За счет этих двух путей регулируется тонус соответствующих мышц при изменениях положения тела.
Все рефлексы спинного мозга делятся на соматические, т.е. двигательные и вегетативные. Соматические рефлексы делятся на сухожильные или миотатические и кожные. Сухожильные рефлексы возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и a-мотонейронов спинного мозга. В результате возникает сокращение мышц, в первую очередь разгибателей. К сухожильным рефлексам относятся коленный, ахиллов, локтевой, кистевой и др., возникающие при механическом раздражении соответствующих сухожилий. Например, коленный является простейшим моносинаптическим, так как в его центральной части только один синапс. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной (объяснение). Спинальные нервные центры находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится.
Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.
(Продолговатый мозг и мост, их функции, участие в регуляции мышечного тонуса)
Продолговатый мозг
Особенности функциональной организации. Продолговатый мозг (medullaoblongata) у человека имеет длину около 25 мм. Он является продолжением спинного мозга. Структурно по разнообразию и строению ядер продолговатый мозг сложнее, чем спинной. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а ядрами к периферии.
В продолговатом мозге находятся оливы, связанные со спинным мозгом, экстрапирамидной системой и мозжечком — это тонкое и клиновидное ядра проприоцептивной чувствительности (ядра Голля и Бурдаха). Здесь же находятся перекресты нисходящих пирамидных путей и восходящих путей, образованных тонким и клиновидным пучками (Голля и Бурдаха), ретикулярная формация.
Продолговатый мозг за счет своих ядерных образований и ретикулярной формации участвует в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов. Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании.
В продолговатом мозге расположены ядра следующих черепных нервов:
пара VIII черепных нервов — преддверно-улитковый нерв состоит из улитковой и преддверной частей. Улитковое ядро лежит в продолговатом мозге;
пара IX — языкоглоточный нерв (п. glossopharyngeus); его ядро образовано 3 частями — двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная — получает информацию от рецепторов вкуса задней трети языка; вегетативная иннервирует слюнные железы;
пара X — блуждающий нерв (n.vagus) имеет 3 ядра: вегетативное иннервирует гортань, пищевод, сердце, желудок, кишечник, пищеварительные железы; чувствительное получает информацию от рецепторов альвеол легких и других внутренних органов и двигательное (так называемое обоюдное) обеспечивает последовательность сокращения мышц глотки, гортани при глотании;
пара XI — добавочный нерв (n.accessorius); его ядро частично расположено в продолговатом мозге;
пара XII — подъязычный нерв (n.hypoglossus) является двигательным нервом языка, его ядро большей частью расположено в продолговатом мозге.
Сенсорные функции. Продолговатый мозг регулирует ряд сенсорных функций: рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; рецепцию слуховых раздражений — в ядре улиткового нерва; рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолготоватого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Проводниковые функции. Через продолготоватый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Рефлекторные функции. Многочисленные рефлексы продолговатого мозга делят на жизненно важные и нежизненно важные, однако такое представление достаточно условно. Дыхательные и сосудодвигательные центры продолговатого мозга можно отнести к жизненно важным центрам, так как в них замыкается ряд сердечных и дыхательных рефлексов.
Продолговатый мозг организует и реализует ряд защитных рефлексов: рвоты, чиханья, кашля, слезоотделения, смыкания век. Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга, отсюда идет команда к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате реализуется тот или иной защитный рефлекс. Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания.
Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо.
Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы, соответствующей моменту прямолинейного или вращательного движения.
Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов.
Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, сужается просвет бронхов.
Деятельность ядер блуждающего нерва проявляется также в усилении секреции бронхиальных, желудочных, кишечных желез, в возбуждении поджелудочной железы, секреторных клеток печени.
В продолговатом мозге локализуется центр слюноотделения, парасимпатическая часть которого обеспечивает усиление общей секреции, а симпатическая — белковой секреции слюнных желез.
В структуре ретикулярной формации продолговатого мозга расположены дыхательный и сосудодвигательный центры. Особенность этих центров в том, что их нейроны способны возбуждаться рефлекторно и под действием химических раздражителей.
Дыхательный центр локализуется в медиальной части ретикулярной формации каждой симметричной половины продолговатого мозга и разделен на две части, вдоха и выдоха.
В ретикулярной формации продолговатого мозга представлен другой жизненно важный центр — сосудодвигательный центр (регуляции сосудистого тонуса). Он функционирует совместно с вышележащими структурами мозга и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др. Это обусловлено тем, что ретикулярная формация продолговатого мозга имеет синаптические связи с гипоталамусом и другими центрами.
В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного мозга. На дне IV желудочка расположены нейроны «голубого пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса.
Симптомы повреждения. Повреждение левой или правой половины продолговато мозга выше перекреста восходящих путей проприоцептивной чувствительности вызывает на стороне повреждения нарушения чувствительности и работы мышц лица и головы. В то же время на противоположной стороне относительно стороны повреждения наблюдаются нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это объясняется тем, что восходящие и нисходящие проводящие пути из спинного мозга и в спинной мозг перекрещиваются, а ядра черепных нервов иннервируют свою половину головы, т. е. черепные нервы не перекрещиваются.
Мост
Мост (ponscerebri, ponsVarolii) располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции.
В состав моста входят ядра лицевого, тройничного, отводящего, преддверно-улиткового нерва (вестибулярные и улитковые ядра), ядра преддверной части преддверно-улиткового нерва (вестибулярного нерва): латеральное (Дейтерса) и верхнее (Бехтерева). Ретикулярная формация моста тесно связана с ретикулярной формацией среднего и продолговатого мозга.
Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка.
Сенсорные функции моста обеспечиваются ядрами преддверно-улиткового, тройничного нервов. Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная часть преддверно-улиткового нерва — в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
Чувствительное ядро тройничного нерва получает сигналы от рецепторов кожи лица, передних отделов волосистой части головы, слизистой оболочки носа и рта, зубов и конъюнктивы глазного яблока. Лицевой нерв (п. facialis) иннервирует все мимические мышцы лица. Отводящий нерв (п. abducens) иннервирует прямую латеральную мышцу, отводящую глазное яблоко кнаружи.
Двигательная порция ядра тройничного нерва (п. trigeminus) иннервирует жевательные мышцы, мышцу, натягивающую барабанную перепонку, и мышцу, натягивающую небную занавеску.
Проводящая функция моста. Обеспечивается продольно и поперечно расположенными волокнами. Поперечно расположенные волокна образуют верхний и нижний слои, а между ними проходят идущие из коры большого мозга пирамидные пути. Между поперечными волокнами расположены нейронные скопления — ядра моста. От их нейронов начинаются поперечные волокна, которые идут на противоположную сторону моста, образуя среднюю ножку мозжечка и заканчиваясь в его коре.
В покрышке моста располагаются продольно идущие пучки волокон медиальной петли (lemniscusmedialis). Они пересекаются поперечно идущими волокнами трапециевидного тела (corpustrapezoideum), представляющие собой аксоны улитковой части преддверно-улиткового нерва противоположной стороны, которые заканчиваются в ядре верхней оливы (olivasuperior). От этого ядра идут пути боковой петли (lemniscuslateralis), которые направляются в заднее четверохолмие среднего мозга и в медиальные коленчатые тела промежуточного мозга.
В покрышке мозга локализуются переднее и заднее ядра трапециевидного тела и латеральной петли. Эти ядра вместе с верхней оливой обеспечивают первичный анализ информации от органа слуха и затем передают информацию в задние бугры четверохолмий.
В покрышке также расположены длинный медиальный и тектоспинальный пути.
Собственные нейроны структуры моста образуют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательной порции ядра и среднее сенсорное ядро тройничного нерва.
Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга и началом этой же системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, в спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга.
Ретикулярная формация моста влияет на кору большого мозга, вызывая ее пробуждение или сонное состояние. В ретикулярной формации моста находятся две группы ядер, которые относятся к общему дыхательному центру. Один центр активирует центр вдоха продолговатого мозга, другой — центр выдоха. Нейроны дыхательного центра, расположенные в мосте, адаптируют работу дыхательных клеток продолговатого мозга в соответствии с меняющимся состоянием организма.
(Средний мозг, его функции, участие в регуляции мышечного тонуса)
Морфофункциональная организация. Средний мозг (mesencephalon) представлен четверохолмием и ножками мозга. Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и ядра черепных (глазодвигательного и блокового) нервов, а также ядра ретикулярной формации.
Сенсорные функции. Реализуются за счет поступления в него зрительной, слуховой информации.
Проводниковая функция. Заключается в том, что через него проходят все восходящие пути к вышележащим таламусу (медиальная петля, спииноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. Это пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функция. Реализуется за счет ядра блокового нерва (n. trochlearis), ядер глазодвигательного нерва (п. oculomotorius), красного ядра (nucleusruber), черного вещества (substantianigra).
Красные ядра располагаются в верхней части ножек мозга. Они связаны с корой большого мозга (нисходящие от коры пути), подкорковыми ядрами, мозжечком, спинным мозгом (красноядерно-спинномозговой путь). Базальные ганглии головного мозга, мозжечок имеют свои окончания в красных ядрах. Нарушение связей красных ядер с ретикулярной формацией продолговатого мозга ведет к децеребрационной ригидности. Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрационной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. Это влияние максимально в отсутствие тормозных влияний красного ядра и вышележащих структур, а также мозжечка. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрационная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры большого мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному тракту и тем самым регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению.
Другое функционально важное ядро среднего мозга — черное вещество — располагается в ножках мозга, регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти руки, например при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который поставляется аксональным транспортом к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц. Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом. В то же время при длительном удержании определенной позы происходят пластические изменения в мышцах за счет изменения их коллоидных свойств, что обеспечивает наименьшие затраты энергии. Регуляция этого процесса осуществляется клетками черного вещества.
Нейроны ядер глазодвигательного и блокового нервов регулируют движение глаза вверх, вниз, наружу, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика.
Рефлекторные функции. Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора (вместе с латеральными коленчатыми телами промежуточного мозга), нижние — слухового (вместе с медиальными коленчатыми телами промежуточного мозга). В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными. В последнем случае они реагируют только на один признак раздражения, например смену света и темноты, направление движения светового источника и т. д. Основная функция бугров четверохолмия — организация реакции настораживания и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные или звуковые сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы.
У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство.
При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
Ретикулярная формация ствола мозга
Ретикулярная формация (formatioreticularis; РФ) мозга представлена сетью нейронов с многочисленными диффузными связями между собой и практически со всеми структурами центральной нервной системы. РФ располагается в толще серого вещества продолговатого, среднего, промежуточного мозга и изначально связана с РФ спинного мозга. В связи с этим целесообразно ее рассмотреть как единую систему. Сетевые связи нейронов РФ между собой позволили Дейтерсу назвать ее ретикулярной формацией мозга.
РФ имеет прямые и обратные связи с корой большого мозга, базальными ганглиями, промежуточным мозгом, мозжечком, средним, продолговатым и спинным мозгом.
Основной функцией РФ является регуляция уровня активности коры большого мозга, мозжечка, таламуса, спинного мозга.
С одной стороны, генерализованный характер влияния РФ на многие структуры мозга дал основание считать ее неспецифической системой. Однако исследования с раздражением РФ ствола показали, что она может избирательно оказывать активирующее или тормозящее влияние на разные формы поведения, на сенсорные, моторные, висцеральные системы мозга. Сетевое строение обеспечивает высокую надежность функционирования РФ, устойчивость к повреждающим воздействиям, так как локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. С другой стороны, высокая надежность функционирования РФ обеспечивается тем, что раздражение любой из ее частей отражается на активности всей РФ данной структуры за счет диффузности связей.
Большинство нейронов РФ имеет длинные дендриты и короткий аксон. Существуют гигантские нейроны с длинным аксоном, образующие пути из РФ в другие области мозга, например в нисходящем направлении, ретикулоспинальный и руброспинальный. Аксоны нейронов РФ образуют большое число коллатералей и синапсов, которые оканчиваются на нейронах различных отделов мозга. Аксоны нейронов РФ, идущие в кору большого мозга, заканчиваются здесь на дендритах I и II слоев.
Активность нейронов РФ различна и в принципе сходна с активностью нейронов других структур мозга, но среди нейронов РФ имеются такие, которые обладают устойчивой ритмической активностью, не зависящей от приходящих сигналов.
В то же время в РФ среднего мозга и моста имеются нейроны, которые в покое «молчат», т. е. не генерируют импульсы, но возбуждаются при стимуляции зрительных или слуховых рецепторов. Это так называемые специфические нейроны, обеспечивающие быструю реакцию на внезапные, неопознанные сигналы. Значительное число нейронов РФ являются полисенсорными.
В РФ продолговатого, среднего мозга и моста конвергируют сигналы различной сенсорности. На нейроны моста приходят сигналы преимущественно от соматосенсорных систем. Сигналы от зрительной и слуховой сенсорных систем в основном приходят на нейроны РФ среднего мозга.
РФ контролирует передачу сенсорной информации, идущей через ядра таламуса, за счет того, что при интенсивном внешнем раздражении нейроны неспецифических ядер таламуса затормаживаются, тем самым снимается их тормозящее влияние с релейных ядер того же таламуса и облегчается передача сенсорной информации в кору большого мозга.
В РФ моста, продолговатого, среднего мозга имеются нейроны, которые реагируют на болевые раздражения, идущие от мышц или внутренних органов, что создает общее диффузное дискомфортное, не всегда четко локализуемое, болевое ощущение «тупой боли».
Повторение любого вида стимуляции приводит к снижению импульсной активности нейронов РФ, т. е. процессы адаптации (привыкания) присущи и нейронам РФ ствола мозга.
РФ ствола мозга имеет прямое отношение к регуляции мышечного тонуса, поскольку на РФ ствола мозга поступают сигналы от зрительного и вестибулярного анализаторов и мозжечка. От РФ к мотонейронам спинного мозга и ядер черепных нервов поступают сигналы, организующие положение головы, туловища и т. д.
Ретикулярные пути, облегчающие активность моторных систем спинного мозга, берут начало от всех отделов РФ. Пути, идущие от моста, тормозят активность мотонейронов спинного мозга, иннервирующих мышцы-сгибатели, и активируют мотонейроны мышц-разгибателей. Пути, идущие от РФ продолговатого мозга, вызывают противоположные эффекты. Раздражение РФ приводит к тремору, повышению тонуса мышц. После прекращения раздражения вызванный им эффект сохраняется длительно, видимо, за счет циркуляции возбуждения в сети нейронов.
РФ ствола мозга участвует в передаче информации от коры большого мозга, спинного мозга к мозжечку и, наоборот, от мозжечка к этим же системам. Функция данных связей заключается в подготовке и реализации моторики, связанной с привыканием, ориентировочными реакциями, болевыми реакциями, организацией ходьбы, движениями глаз.
Регуляция вегетативной деятельности организма РФ описана в разделе 4.3, здесь же заметим, что наиболее четко эта регуляция проявляется в функционировании дыхательного и сердечно-сосудистых центров. В регуляции вегетативных функций большое значение имеют так называемые стартовые нейроны РФ. Они дают начало циркуляции возбуждения внутри группы нейронов, обеспечивая тонус регулируемых вегетативных систем.
Влияния РФ можно разделить в целом на нисходящие и восходящие. В свою очередь каждое из этих влияний имеет тормозное и возбуждающее действие.
Восходящие влияния РФ на кору большого мозга повышают ее тонус, регулируют возбудимость ее нейронов, не изменяя специфику ответов на адекватные раздражения. РФ влияет на функциональное состояние всех сенсорных областей мозга, следовательно, она имеет значение в интеграции сенсорной информации от разных анализаторов.
РФ имеет прямое отношение к регуляции цикла бодрствование—сон. Стимуляция одних структур РФ приводит к развитию сна, стимуляция других вызывает пробуждение. Г. Мэгун и Д. Моруцци выдвинули концепцию, согласно которой все виды сигналов, идущих от периферических рецепторов, достигают по коллатералям РФ продолговатого мозга и моста, где переключаются на нейроны, дающие восходящие пути в таламус и затем в кору большого мозга.
Возбуждение РФ продолговатого мозга или моста вызывает синхронизацию активности коры большого мозга, появление медленных ритмов в ее электрических показателях, сонное торможение.
Возбуждение РФ среднего мозга вызывает противоположный эффект пробуждения: десинхронизацию электрической активности коры, появление быстрых низкоамплитудных β-подобных ритмов в электроэнцефалограмме.
Г. Бремер (1935) показал, что если перерезать мозг между передними и задними буграми четверохолмия, то животное перестает реагировать на все виды сигналов; если же перерезку произвести между продолговатым и средним мозгом (при этом РФ сохраняет связь с передним мозгом), то животное реагирует на свет, звук и другие сигналы. Следовательно, поддержание активного анализирующего состояния мозга возможно при сохранении связи с передним мозгом.
Реакция активации коры большого мозга наблюдается при раздражении РФ продолговатого, среднего, промежуточного мозга. В то же время раздражение некоторых ядер таламуса приводит к возникновению ограниченных локальных участков возбуждения, а не к общему ее возбуждению, как это бывает при раздражении других отделов РФ.
РФ ствола мозга может оказывать не только возбуждающее, но и тормозное влияние на активность коры мозга.
Нисходящие влияния РФ ствола мозга на регуляторную деятельность спинного мозга были установлены еще И. М. Сеченовым (1862). Им было показано, что при раздражении среднего мозга кристалликами соли у лягушки рефлексы отдергивания лапки возникают медленно, требуют более сильного раздражения или не появляются вообще, т. е. тормозятся.
Г. Мэгун (1945—1950), нанося локальные раздражения на РФ продолговатого мозга, нашел, что при раздражении одних точек тормозятся, становятся вялыми рефлексы сгибания передней лапы, коленный, роговичный. При раздражении РФ в других точках продолговатого мозга эти же рефлексы вызывались легче, были сильнее, т. е. их реализация облегчалась. По мнению Мэгуна, тормозные влияния на рефлексы спинного мозга может оказывать только РФ продолговатого мозга, а облегчающие влияния регулируются всей РФ ствола и спинного мозга.
(Мозжечок, его участие в регуляции моторных и вегетативных функций)
Мозжечок (cerebellum, малый мозг) — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных, непроизвольных движений, в регуляции вегетативных и поведенческих функций.
Особенности морфофункциональной организации и связи мозжечка. Реализация указанных функций обеспечивается следующими морфологическими особенностями мозжечка:
1) кора мозжечка построена достаточно однотипно, имеет стереотипные связи, что создает условия для быстрой обработки информации;
2) основной нейронный элемент коры — клетка Пуркинье, имеет большое количество входов и формирует единственный аксонный выход из мозжечка, коллатерали которого заканчиваются на ядерных его структурах;
3) на клетки Пуркинье проецируются практически все видысенсорных раздражений: проприоцептивные, кожные, зрительные,слуховые, вестибулярные и др.;
4) выходы из мозжечка обеспечивают его связи с корой большого мозга, со стволовыми образованиями и спинным мозгом.
Мозжечок анатомически и функционально делится на старую, древнюю и новую части.
К старой части мозжечка (archicerebellum) — вестибулярный мозжечок — относится клочково-флоккулярная доля. Эта часть имеет наиболее выраженные связи с вестибулярным анализатором, что объясняет значение мозжечка в регуляции равновесия.
Древняя часть мозжечка (paleocerebellum) — спинальный мозжечок — состоит из участков червя и пирамиды мозжечка, язычка, околоклочкового отдела и получает информацию преимущественно от проприорецептивных систем мышц, сухожилий, надкостницы, оболочек суставов.
Новый мозжечок (neocerebellum) включает в себя кору полушарий мозжечка и участки червя; он получает информацию от коры, преимущественно по лобно-мостомозжечковому пути, от зрительных и слуховых рецептирующих систем, что свидетельствует об его участии в анализе зрительных, слуховых сигналов и организации на них реакции.
Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Верхний (I) слой коры мозжечка — молекулярный слой, состоит из параллельных волокон, разветвлений дендритов и аксонов II и III слоев. В нижней части молекулярного слоя встречаются корзинчатые и звездчатые клетки, которые обеспечивают взаимодействие клеток Пуркинье.
Средний (II) слой коры образован клетками Пуркинье, выстроенными в один ряд и имеющими самую мощную в ЦНС дендритную систему. На дендритном поле одной клетки Пуркинье может быть до 60 000 синапсов. Следовательно, эти клетки выполняют задачу сбора, обработки и передачи информации. Аксоны клеток Пуркинье являются единственным путем, с помощью которого кора мозжечка передает информацию в его ядра и ядра структуры большого мозга.
Под II слоем коры (под клетками Пуркинье) лежит гранулярный (III) слой, состоящий из клеток-зерен, число которых достигает 10 млрд. Аксоны этих клеток поднимаются вверх, Т-образно делятся на поверхности коры, образуя дорожки контактов с клетками Пуркинье. Здесь же лежат клетки Гольджи.
Из мозжечка информация уходит через верхние и нижние ножки. Через верхние ножки сигналы идут в таламус, в мост, красное ядро, ядра ствола мозга, в ретикулярную формацию среднего мозга. Через нижние ножки мозжечка сигналы идут в продолговатый мозг к его вестибулярным ядрам, оливам, ретикулярной формации. Средние ножки мозжечка связывают новый мозжечок с лобной долей мозга.
Импульсная активность нейронов регистрируется в слое клеток Пуркинье и гранулярном слое, причем частота генерации импульсов этих клеток колеблется от 20 до 200 в секунду. Клетки ядер мозжечка генерируют импульсы значительно реже — 1—3 импульса в секунду.
Стимуляция верхнего слоя коры мозжечка приводит к длительному (до 200 мс) торможению активности клеток Пуркинье. Такое же их торможение возникает при световых и звуковых сигналах. Суммарные изменения электрической активности коры мозжечка на раздражение чувствительного нерва любой мышцы выглядят в форме позитивного колебания (торможение активности коры, гиперполяризация клеток Пуркинье), которое наступает через 15— 20 мс и длится 20—30 мс, после чего возникает волна возбуждения, длящаяся до 500 мс (деполяризация клеток Пуркинье).
В кору мозжечка от кожных рецепторов, мышц, суставных оболочек, надкостницы сигналы поступают по так называемым спинно-мозжечковым трактам: по заднему (дорсальному) и переднему (вентральному). Эти пути к мозжечку проходят через нижнюю оливу продолговатого мозга. От клеток олив идут так называемые лазающие волокна, которые ветвятся на дендритах клеток Пуркинье.
Ядра моста посылают афферентные пути в мозжечок, образующие мшистые волокна, которые оканчиваются на клетках-зернах III слоя коры мозжечка. Между мозжечком и голубоватым местом среднего мозга существует афферентная связь с помощью адренергических волокон. Эти волокна способны диффузно выбрасывать норадреналин в межклеточное пространство коры мозжечка, тем самым гуморально изменяют состояние возбудимости его клеток.
Аксоны клеток III слоя коры мозжечка вызывают торможение клеток Пуркинье и клеток-зерен своего же слоя.
Клетки Пуркинье в свою очередь тормозят активность нейронов ядер мозжечка. Ядра мозжечка имеют высокую тоническую активность и регулируют тонус ряда моторных центров промежуточного, среднего, продолговатого, спинного мозга.
Подкорковая система мозжечка состоит из трех функционально разных ядерных образований: ядра шатра, пробковидного, шаровидного и зубчатого ядра.
Ядро шатра получает информацию от медиальной зоны коры мозжечка и связано с ядром Дейтерса и РФ продолговатого и среднего мозга. Отсюда сигналы идут по ретикулоспинальному пути к мотонейронам спинного мозга.
Промежуточная кора мозжечка проецируется на пробковидное и шаровидное ядра. От них связи идут в средний мозг к красному ядру, далее в спинной мозг по руброспинальному пути. Второй путь от промежуточного ядра идет к таламусу и далее в двигательную зону коры большого мозга.
Зубчатое ядро, получая информацию от латеральной зоны коры мозжечка, связано с таламусом, а через него — с моторной зоной коры большого мозга.
Мозжечковый контроль двигательной активности. Эфферентные сигналы из мозжечка к спинному мозгу регулируют силу мышечных сокращений, обеспечивают способность к длительному тоническому сокращению мышц, способность сохранять оптимальный тонус мышц в покое или при движениях, соразмерять произвольные движения с целью этого движения, быстро переходить от сгибания к разгибанию и наоборот.
Мозжечок обеспечивает синергию сокращений разных мышц при сложных движениях. Например, делая шаг при ходьбе, человек заносит вперед ногу, одновременно центр тяжести туловища переносится вперед при участии мышц спины. В тех случаях, когда мозжечок не выполняет своей регуляторной функции, у человека наблюдаются расстройства двигательных функций, что выражается следующими симптомами.
1) астения (astenia — слабость) — снижение силы мышечного сокращения, быстрая утомляемость мышц;
2) астазия (astasia, от греч.а — не, stasia — стояние) — утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение и т. д.;
3) дистония (distonia — нарушение тонуса) — непроизвольное повышение или понижение тонуса мышц;
4) тремор (tremor — дрожание) — дрожание пальцев рук, кистей, головы в покое; этот тремор усиливается при движении;
5) дисметрия (dismetria — нарушение меры) — расстройство равномерности движений, выражающееся либо в излишнем, либо недостаточном движении. Больной пытается взять предмет со стола и проносит руку за предмет (гиперметрия) или не доносит ее до предмета (гипометрия);
6) атаксия (ataksia, от греч.а — отрицание, taksia — порядок) — нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности. Проявлениями атаксии являются так же адиадохокинез, асинергия, пьяная-шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз—вверх. При асинергии мышц он не способен сесть из положения лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы. Врожденных двигательных актов у человека не так уж много (например, сосание), большинство же движений он выучивает в течение жизни и они становятся автоматическими (ходьба, письмо и т.д.). Когда нарушается функция мозжечка, движения становятся неточными, негармоничными, разбросанными, часто не достигают цели.
Данные о том, что повреждение мозжечка ведет к расстройствам движений, которые были приобретены человеком в результате обучения, позволяют сделать вывод, что само обучение шло с участием мозжечковых структур, а следовательно, мозжечок принимает участие в организации процессов высшей нервной деятельности;
7) дизартрия (disartria) — расстройство организации речевой моторики. При повреждении мозжечка речь больного становится растянутой, слова иногда произносятся как бы толчками (скандированная речь).
При повреждении мозжечка наблюдается повышение тонуса мышц-разгибателей. Регуляция мышечного тонуса с помощью мозжечка происходит следующим образом: проприоцептивные сигналы о тонусе мышц поступают в область червя и клочково-узелковую долю, отсюда — в ядро шатра, далее — к ядру преддверия и РФ продолговатого и среднего мозга и, наконец, по ретикулярно- и вестибулоспинальным путям к нейронам передних рогов спинного мозга, иннервирующих мышцы, от которых поступили сигналы. Следовательно, регуляция мышечного тонуса реализуется по принципу обратной связи.
Следует отметить, что характер влияния на тонус мышц определяется частотой генерации импульсов нейронов ядра шатра. При высокой частоте (30—300 имп/с) тонус мышц-разгибателей снижается, при низкой (2—10 имп/с) — увеличивается.
Промежуточная область коры мозжечка получает информацию по спинальным трактам от двигательной области коры большого мозга (прецентральной извилины), по коллатералям пирамидного пути, идущего в спинной мозг. Коллатерали заходят в мост, а оттуда — в кору мозжечка. Следовательно, за счет коллатералей мозжечок получает информацию о готовящемся произвольном движении, и возможность участвовать в обеспечении тонуса мышц, необходимого для реализации этого движения.
Латеральная кора мозжечка получает информацию из двигательной области коры большого мозга. В свою очередь латеральная кора посылает информацию в зубчатое ядро мозжечка, отсюда по мозжечково-кортикальному пути в сенсомоторную область коры большого мозга (постцентральная извилина), а через мозжечково-рубральный путь к красному ядру и от него по руброспинальному пути к передним рогам спинного мозга. Параллельно сигналы по пирамидному пути идут к тем же передним рогам спинного мозга.
Таким образом, мозжечок, получив информацию о готовящемся движении, корректирует программу подготовки этого движения в коре и одновременно готовит тонус мускулатуры для реализации этого движения через спинной мозг.
Изменение тонуса мышц после повреждения мозжечка обусловлено тем, что исчезает торможение лабиринтных и миотатических рефлексов, которое в норме осуществляется мозжечком. В норме вестибулярные ядра активируют мотонейроны спинного мозга мышц-разгибателей, а мозжечок тормозит активность нейронов преддверного ядра. При повреждении мозжечка вестибулярные ядра бесконтрольно активируют мотонейроны передних рогов спинного мозга, в результате повышается тонус мышц-разгибателей конечностей.
При повреждении мозжечка усиливаются и проприоцептивные рефлексы спинного мозга (рефлексы, вызываемые при раздражении рецепторов сухожилий, мышц, надкостницы, оболочек суставов), но в этом случае снимается тормозное влияние на мотонейроны спинного мозга ретикулярной формации продолговатого мозга.
В норме мозжечок активирует пирамидные нейроны коры большого мозга, которые тормозят активность мотонейронов спинного мозга. Чем больше мозжечок активирует пирамидные нейроны коры, тем более выражено торможение мотонейронов спинного мозга. При повреждении мозжечка это торможение исчезает, так как активация пирамидных клеток прекращается.
Таким образом, в случае повреждения мозжечка активируются нейроны вестибулярных ядер и ретикулярной формации продолговатого мозга, которые активируют мотонейроны спинного мозга. Одновременно активность пирамидных нейронов снижается, а следовательно, снижается их тормозное влияние на те же мотонейроны спинного мозга. В итоге, получая возбуждающие сигналы от продолговатого мозга при одновременном уменьшении тормозных влияний от коры большого мозга (после повреждения структур мозжечка), мотонейроны спинного мозга активируются и вызывают гипертонус мышц.
Взаимодействие мозжечка и коры большого мозга. Это взаимодействие организовано соматотопически. Функционально мозжечок может оказывать облегчающее, тормозящее и компенсаторное влияние на реализацию функций коры большого мозга.
Роль взаимодействия лобной доли коры большого мозга с мозжечком хорошо проявляется при частичных повреждениях мозжечка. Одномоментное удаление мозжечка приводит к гибели человека, в то же время, если удаляется часть мозжечка, это вмешательство, как правило, не смертельно. После операции частичного удаления мозжечка возникают симптомы его повреждения (тремор, атаксия, астения и т. д.), которые затем исчезают. Если на фоне исчезновения мозжечковых симптомов нарушается функция лобных долей мозга, то мозжечковые симптомы возникают вновь. Следовательно, кора лобных долей большого мозга компенсирует расстройства, вызываемые повреждением мозжечка. Механизм данной компенсации реализуется через лобно-мостомозжечковый тракт.
Мозжечок за счет своего влияния на сенсомоторную область коры может изменять уровень тактильной, температурной, зрительной чувствительности. Оказалось, что повреждение мозжечка снижает уровень восприятия критической частоты мельканий света (наименьшая частота мельканий, при которой световые стимулы воспринимаются не как отдельные вспышки, а как непрерывный свет).
Удаление мозжечка приводит к ослаблению силы процессов возбуждения и торможения, нарушению баланса между ними, развитию инертности. Выработка двигательных условных рефлексов после удаления мозжечка затрудняется, особенно в случаях формирования локальной, изолированной двигательной реакции. Точно так же замедляется выработка пищевых условных рефлексов, увеличивается скрытый (латентный) период их вызова.
Влияние мозжечка на вегетативные функции. Мозжечок оказывает угнетающее и стимулирующее влияние на работу сердечнососудистой, дыхательной, пищеварительной и других систем организма. В результате двойственного влияния мозжечок стабилизирует, оптимизирует функции систем организма.
Сердечно-сосудистая система реагирует на раздражение мозжечка либо усилением (например, прессорные рефлексы), либо снижением этой реакции. Направленность реакции зависит от фона, на котором она вызывается. При раздражении мозжечка высокое кровяное давление снижается, а исходное низкое — повышается. Раздражение мозжечка на фоне учащенного дыхания (гиперпноэ) снижает частоту дыхания. При этом одностороннее раздражение мозжечка вызывает на своей стороне снижение, а на противоположной — повышение тонуса дыхательных мышц.
Удаление или повреждение мозжечка приводит к уменьшению тонуса мускулатуры кишечника, из-за низкого тонуса нарушается эвакуация содержимого желудка и кишечника. Нарушается также нормальная динамика секреции и всасывания в желудке и кишечнике.
Обменные процессы при повреждении мозжечка идут более интенсивно, гипергликемическая реакция (увеличение количества глюкозы в крови) на введение глюкозы в кровь или на прием ее с пищей возрастает и сохраняется дольше, чем в норме, ухудшается аппетит, наблюдается исхудание, замедляется заживление ран, волокна скелетных мышц подвергаются жировому перерождению.
При повреждении мозжечка нарушается генеративная функция, что проявляется в нарушении последовательности процессов родовой деятельности. При возбуждении или повреждении мозжечка мышечные сокращения, сосудистый тонус, обмен веществ и т. д. реагируют так же, как при активации или повреждении симпатического отдела вегетативной нервной системы.
Таким образом, мозжечок принимает участие в различных видах деятельности организма: моторной, соматической, вегетативной, сенсорной, интегративной и т. д. Однако эти функции мозжечок реализует через другие структуры центральной нервной системы. Мозжечок выполняет функцию оптимизации отношений между различными отделами нервной системы, что реализуется, с одной стороны, активацией отдельных центров, с другой — удержанием этой активности в определенных рамках возбуждения, лабильности и т. д. После частичного повреждения мозжечка могут сохраняться все функции организма, но сами функции, порядок их реализации, количественное соответствие потребностям трофики организма нарушаются.
(Таламус, функциональные особенности, значение его ядер)
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
(Гипоталамус, функциональные особенности, значение его ядер)
Гипоталамус (hypothalamus, подбугорье) — структура промежуточного мозга, входящая в лимбическую систему, организующая эмоциональные, поведенческие, гомеостатические реакции организма.
Морфофункциональная организация. Гипоталамус имеет большое число нервных связей с корой большого мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом.
В состав гипоталамуса входят серый бугор, воронка с нейрогипофизом и сосцевидные тела. Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 2) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра; 5) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра.
Ядра гипоталамуса имеют мощное кровоснабжение, подтверждением чему служит тот факт, что ряд ядер гипоталамуса обладает изолированным дублирующим кровоснабжением из сосудов артериального круга большого мозга (виллизиев круг). На 1 мм2 площади гипоталамуса приходится до 2600 капилляров, в то время как на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе — 350, в бледном шаре — 550, в затылочной доле коры большого мозга (зрительной коре) — 900. Капилляры гипоталамуса высокопроницаемы для крупномолекулярных белковых соединений, к которым относятся нуклеопротеиды, что объясняет высокую чувствительность гипоталамуса к нейровирусным инфекциям, интоксикациям, гуморальным сдвигам.
У человека гипоталамус окончательно созревает к возрасту 13— 14 лет, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. За счет мощных афферентных связей с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, корой большого мозга гипоталамус получает информацию о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола мозга и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые и определяют специфику функций самого гипоталамуса. К этим особенностям относятся чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреции пептидов, нейромедиаторов и др.
Роль гипоталамуса в регуляции вегетативных функций. Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
Раздражение ядер передней группы сопровождается парасимпатическими эффектами. Раздражение ядер задней группы вызывает симпатические эффекты в работе органов. Стимуляция ядер средней группы приводит к снижению влияний симпатического отдела автономной нервной системы. Указанное распределение функций гипоталамуса не абсолютно. Все структуры гипоталамуса способны в разной степени вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсирующие отношения.
В целом за счет большого количества связей, полифункциональности структур гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций. Так, в гипоталамусе располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости, регуляции цикла бодрствование—сон. Все эти центры реализуют свои функции путем активации или торможеиия автономного (вегетативного) отдела нервной системы, эндокринной системы, структур ствола и переднего мозга. Нейроны ядер передней группы гипоталамуса продуцируют вазопрессин, или антидиуретический гормон (АДГ), окситоцин и другие пептиды, которые по аксонам попадают в заднюю долю гипофиза — нейрогипофиз.
Нейроны ядер срединной группы гипоталамуса продуцируют так называемые рилизинг-факторы (либерины) и ингибирующие факторы (статины), которые регулируют активность передней доли гипофиза — аденогипофиз. В нем образуются такие вещества, как соматотропный, тиреотропный и другие гормоны (см. раздел 5.2.2). Наличие такого набора пептидов в структурах гипоталамуса свидетельствует о присущей им нейросекреторной функции.
Они также обладают детектирующей функцией: реагируют на изменения температуры крови, электролитного состава и осмотического давления плазмы, количества и состав гормонов крови.
Олдс (Olds) описал поведение крыс, которым вживляли электроды в ядра гипоталамуса и давали возможность самостоятельно стимулировать эти ядра. Оказалось, что стимуляция некоторых ядер приводила к негативной реакции. Животные после однократной самостимуляции больше не подходили к педали, замыкающей стимулирующий ток. При самостимуляции других ядер животные нажимали на педаль часами, не обращая внимания на пищу, воду и др.
Исследования Дельгадо (Delgado) во время хирургических операций показали, что у человека раздражение аналогичных участков вызывало эйфорию, эротические переживания. В клинике показано также, что патологические процессы в гипоталамусе могут сопровождаться ускорением полового созревания, нарушением менструального цикла, половой функции.
Раздражение передних отделов гипоталамуса может вызывать у животных пассивно-оборонительную реакцию, ярость страх, а раздражение заднего гипоталамуса вызывает активную агрессию.
Раздражение заднего гипоталамуса приводит к экзофтальму, расширению зрачков, повышению кровяного давления, сужению просвета артериальных сосудов, сокращениям желчного, мочевого пузырей. Могут возникать взрывы ярости с описанными симпатическими проявлениями. Уколы в области гипоталамуса вызывают глюкозурию, полиурию. В ряде случаев раздражение вызывало нарушение теплорегуляции: животные становились пойкилотермными, у них не возникало лихорадочное состояние.
Гипоталамус является также центром регуляции цикла бодрствование — сон. При этом задний гипоталамус активизирует бодрствование, стимуляция переднего вызывает сон. Повреждение заднего гипоталамуса может вызвать так называемый летаргический сон.
Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза (см. раздел 5.2.2).
В гипоталамусе и гипофизе образуются также нейрорегуляторные пептиды — энкефалины, эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса и т. д.
(Лимбическая система мозга)
Лимбическая система представляет собой функциональное объединение структур мозга, участвующих в организации эмоционально-мотивационного поведения, таких как пищевой, половой, оборонительный инстинкты. Эта система участвует в организации цикла бодрствование—сон.
Лимбическая система как филогенетически древнее образование оказывает регулирующее влияние на кору большого мозга и подкорковые структуры, устанавливая необходимое соответствие уровней их активности.
Морфофункциональная организация. Структуры лимбической системы включают в себя 3 комплекса. Первый комплекс — древняя кора (препериформная, периамигдалярная, диагональная кора), обонятельные луковицы, обонятельный бугорок, прозрачная перегородка (рис. 4.12).
Вторым комплексом структур лимбической системы является старая кора, куда входят гиппокамп, зубчатая фасция, поясная извилина.
Третий комплекс лимбической системы — структуры островковой коры, парагиппокамповая извилина.
И, наконец, в лимбическую систему включают подкорковые структуры: миндалевидные тела, ядра прозрачной перегородки, переднее таламическое ядро, сосцевидные тела.
Особенностью лимбической системы является то, что между ее структурами имеются простые двусторонние связи и сложные пути, образующие множество замкнутых кругов (см. рис. 4.12). Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и тем самым для сохранения в ней единого состояния и навязывание этого состояния другим системам мозга.
В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную специфику. К ним относится круг Пейпеса (гиппокамп à сосцевидные тела à передние ядра таламуса à кора поясной извилины à парагиппокампова извилина à гиппокамп). Этот круг имеет отношение к памяти и процессам обучения.
Другой круг (миндалевидное тело à гипоталамус à мезенцефальные структуры à миндалевидное тело) регулирует агрессивно-оборонительные, пищевые и сексуальные формы поведения.
Считается, что образная (иконическая) память формируется кортико-лимбико-таламо-кортикальным кругом. Круги разного функционального назначения связывают лимбическую систему со многими структурами центральной нервной системы, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой.
Например, включение хвостатого ядра в один из кругов лимбической системы определяет ее участие в организации тормозных процессов высшей нервной деятельности.
Большое количество связей в лимбической системе, своеобразное круговое взаимодействие ее структур создают благоприятные условия для реверберации возбуждения по коротким и длинным кругам. Это, с одной стороны, обеспечивает функциональное взаимодействие частей лимбической системы, с другой — создает условия для запоминания. Обилие связей лимбической системы со структурами центральной нервной системы затрудняет выделение функций мозга, в которых она не принимала бы участия. Так, лимбическая система имеет отношение к регулированию уровня реакции автономной, соматической систем при эмоционально-мотивационной деятельности, регулированию уровня внимания, восприятия, воспроизведения эмоционально значимой информации. Лимбическая система определяет выбор и реализацию адаптационных форм поведения, динамику врожденных форм поведения, поддержание гомеостаза, генеративных процессов. Наконец, она обеспечивает создание эмоционального фона, формирование и реализацию процессов высшей нервной деятельности.
Нужно отметить, что древняя и старая кора лимбической системы имеет прямое отношение к обонятельной функции. В свою очередь обонятельный анализатор, как самый древний из анализаторов, является неспецифическим активатором всех видов деятельности коры большого мозга.
Некоторые авторы называют лимбическую систему висцеральным мозгом, т. е. структурой ЦНС, участвующей в регуляции деятельности внутренних органов. И действительно, миндалевидные тела, прозрачная перегородка, обонятельный мозг при их возбуждении изменяют активность вегетативных систем организма в соответствии с условиями окружающей среды. Это стало возможно благодаря установлению морфологических и функциональных связей с более молодыми образованиями мозга, обеспечивающими взаимодействие экстероцептивных, интероцептивных систем и коры височной доли.
Наиболее полифункциональными образованиями лимбической системы являются гиппокамп и миндалевидные тела. Физиология этих структур наиболее изучена.
(Парность в деятельности коры больших полушарий.Функциональная асимметрия коры у человека)
В проблеме соотношения задатков и способностей не был рассмотрен вопрос о специальных задатках и способностях. Среди огромного множества разновидностей человеческой деятельности есть такие, которыми может овладеть каждый индивид с нормой общего интеллекта, а есть и такие, которыми любой овладеть не может. Даже высокий интеллектуальный потенциал не может обеспечить успешность, скорость и легкость овладения некоторыми видами деятельности. В таких случаях психологи говорят об отсутствии у личности специальных способностей и стараются выяснить причины такой невозможности.
Рядом с общими способностями выделяют специальные человеческие способности, понимая под ними такие психологические особенности, которые присущи одним индивидам, и отсутствуют у других. К специальным способностям относятся музыкальные, художественные, математические, литературные, поэтические и прочие способности. Действительно, не каждый из нас может стать художником или поэтом, музыкантом или математиком (имеется в виду - способным) даже при использовании совершеннейших методов для развития специальных способностей. Исходя из соотношения "врожденно - приобретенное", будем учитывать, что задатки являются необходимым условием для развития способностей, а природа задатков скрыта во врожденных конструктивных и функциональных особенностях головного мозга человека.
В исследованиях И. П. Павлова и его сотрудников были установлены общие типы ВНД, одинаковые для животных и людей. Стараясь объяснить отличие между высшей нервной деятельностью человека и животного, Павлов подчеркивал, что в психике человека имеет место "чрезвычайное приложение" - речь. Это прибавление Павлов назвал второй сигнальной системой в отличие от первой сигнальной системы (образы ощущений и восприятия). Великий физиолог понимал, что в высшей нервной деятельности человека существует относительное преобладание одной сигнальной системы над другой. С его точки зрения по преобладанию люди могут быть разделены на три специальных типа: "художественный", "мыслительный" и "средний". Изобразим такое преобладание схематично:
ІСС > ІІСС - "художник"
ІСС = ІІСС - "средний"
ІСС < ІІСС - "мыслитель"
Последователи Павловского учения о двух сигнальных системах начали изучать условные "рефлексы рефлексов", то есть образование у людей условных рефлексов не только на первосигнальный раздражитель (свет, цвет, звук, удар током), а и на слово, которое означает раздражитель. Такие исследования проводились, как физиологами, так и психологами. Кстати, исследование условных рефлексов на речевые раздражители велись и на кафедре психологии Одесского университета. Но результаты этих исследований мало что давали для дифференциации специальных задатков и способностей. Ученым понадобилось еще пол столетия, чтобы понять чем же действительно обусловлена детерминация специальных способностей. В конце XІХ и в начале XX столетия, встречаясь с фактами поражений больших полушарий головного мозга человека, исследователи обнаружили разные варианты нарушений и расстройств отдельных свойств высших психических функций (восприятия, памяти, речи, мышления, интеллекта, личностных свойств). Эти факты разрешили сделать вывод, что определенные участки коры больших полушарий головного мозга имеют свое специфическое назначение и функционально обеспечивают соответствующие составные психики в интегративной деятельности личности. Накопление фактов показало, что в левом полушарии сосредоточены центры речи, а правое полушарие обеспечивает образное отражение объектов окружающего мира. Направление таких исследований получило название "нейропсихология". Одним из основателей этого направления был профессор Лурия А. Р.
(Большой вклад в понимание функций правого и левого полушарий внесли наблюдение за пациентами в условиях нейрохирургического вмешательства. Для проведения операций использовался метод торможения функций одного из полушарий головного мозга. При заторможенности одного полушария, второе полушарие начинало вести себя в режиме максимума собственных функциональных возможностей. Ученые приходили к выводу, что в каждой личности находятся два человека: левополушарный и правополушарный. Так, левополушарный человек очень говорлив, находится в эйфоричном состоянии положительных эмоций, строит нереальные "прожекты", целенаправлен в будущее. Правополушарный человек, наоборот, молчалив, агрессивен, связан с образами прошлого. Как очень точно подметили известные исследователи функций правого и левого полушарий головного мозга человека Н. М. Брагина и Т. А. Доброхотова, личность переживает собственное время, которое "течет" слева направо. Здесь имеется в виду функционально асимметрическая ориентация левого полушария в будущее, а правого в прошлое.
Во многих экспериментальных исследованиях было выяснено, что восприятие предметов окружающего мира обеспечивается именно парной работой анализаторов (бинокулярное зрение, бинауральный слух, бимануальное ощупывание).
Каждый анализатор, как выяснилось, имеет свое центральное представительство в коре больших полушарий головного мозга. Так, левый глаз, левое ухо, левая рука имеют свои представительства в правом полушарии, а правый глаз, правое ухо, правая рука - в левом полушарии. В парной работе один из анализаторов выполняет функцию ведущего, а второй ведомого. Соответственно с представительством анализаторов можно говорить о ведущем и ведомом полушариях головного мозга. Эти факты показывают, что левое и правое полушария функционально неравнозначны в обеспечении отображения предметной среды. Соответственно с этой функциональной неравнозначностью относительно медиальной плоскости пространство человека делится на левое и правое.
В других исследованиях было установлено, что преобладание функций правого полушария над левым обуславливает леворукость, а преобладание функций левого полушария над правым обуславливает праворукость} Так, изучая преобладания функций органов в парной работе анализаторов (ведущий - ведомый орган) были разработаны исследовательские приемы функционального преобладания левого - правого полушарий головного мозга человека. Объединяя все признаки преобладаний, ученые стали говорить о функциональных асимметриях больших полушарий головного мозга. Было также установлено, что преобладание может быть полным, или частичным (парциальным). Учитывая все накопленные в этом направлении факты, психологи пришли к выводу, что именно функциональные асимметрии мозга могут рассматриваться как задатки специальных способностей индивида.
Мы рассматриваем задатки как врожденные особенности строения головного мозга. Исследования в этом направлении показывают, что каждый индивид рождается полушарно асимметричным. Так, по данным некоторых исследователей, в человеческой популяции рождается приблизительно одинаковое количество индивидов с преобладанием функций правого полушария (левши) и функций левого полушария (правши). Но, в реальности количество леворуких индивидов встречается значительно реже. Здесь надо сделать некоторые объяснения. Во-первых, имеют место разные варианты разного преобладания функций - полное, или частичное.
Конечно, учитывая сложную структуру коры больших полушарий, вариантов частичного преобладания может быть намного больше, но задача тонкого расщепления этих вариантов преобладания не разрешима в дифференциальной психологии. Во-вторых, левое полушарие, рождаясь "немым", начинает "говорить" (напомним, что все центры речи находятся в левом полушарии) на определенном возрастном периоде развития индивида. В-третьих, так исторически сложилось, что человечество создало праворукую культуру. Все это вместе предопределяет торможение проявления леворукости у определенной части индивидов, которые родились с частичным преобладанием правого полушария над левым. Под влиянием указанных причин эти индивиды превращаются в "скрытыхлеворуких", сохраняя при этом преобладание функций правого полушария над функциями левого. Среди правополушарных под вышеупомянутым социальным давлением "выживает" и сохраняется около половины (по данным американских психологов 20-25%) леворуких индивидов.
Подводя итоги, следует отметить, что в специальной организации психики индивида главную роль играют врожденные функциональные асимметрии больших полушарий головного мозга. Именно последние и определяют задатки, степень проявления которых реализуется в специальных способностях личности. Существуя в единстве, функциональные асимметрии накладывают свою печать на индивидуальные свойства общего типа, прибавляя к формально-динамическим свойствам темперамента специальную направленность.
(Функциональная организация нервных центров, их свойства)
Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров.
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры,
процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное
число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
4. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. Если учесть, что в реальных рефлекторных дугах
имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд.
Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — впоследних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6. Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные стимулы (свет, звук, механические раздражения и т. д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций. Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути» по Ч. Шеррингтону.
9. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
В доминантном очаге устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данныхусловий ритм работы, когда этот очаг становится наиболее чувствительным. Доминирующее значение такого очага (нервного центра) определяет его угнетающее влияние на другие соседние очаги возбуждения. Доминантный очаг возбуждения «притягивает» к себе возбуждение других возбужденных зон (нервных центров). Принцип доминанты определяет формирование главенствующего (активирующего) возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы. При всей сложности складывающихся отношений между старыми, древними и эволюционно новыми нервными образованиями стволовой части мозга общая схема взаимных влияний может быть представлена следующим образом: восходящие влияния (от нижележащих «старых» нервных структур к вышележащим «новым» образованиям) преимущественно носят возбуждающий стимулирующий характер, нисходящие (от вышележащих «новых» нервных образований к нижележащим «старым» нервным структурам) носят угнетающий тормозной характер. Эта схема согласуется с представлением о росте в процессе эволюции роли и значения тормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
(Особенности распространения возбуждения в центральной нервной системе)
Рассматривая основные принципы и особенности распространения возбуждения в ЦНС, а по сути, и свойства нервных центров, можно выделить следующие моменты.
1. Одностороннее проведение возбуждения. В ЦНС - в ее центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении, например, от афферентного нейрона к эфферентному, а не наоборот. Обусловлено это особенностями расположения и характером функционирования химического синапса.
2. Суммация возбуждений (аналогично можно говорить и о суммации торможения). На нейроне в области его аксонального холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона. Если с определенным интервалом к нейрону в точку А приходят импульсы, они вызывают генерацию в этой области ВПСП. Если эти ВПСП не достигают критического уровня деполяризации, то потенциал действия не возникает. Если же частота следования достаточно большая, то происходит в этом месте суммация ВПСП, при достижении ВПСП критического уровня деполяризации возникает ПД, нейрон возбуждается. Это явление носит название временной суммации (происходит суммация следов возбуждения во времени). В ЦНС имеет место и пространственная суммация: возбуждения, приходящие в точку В, А, С нейрона (даже если они сами по себе - подпороговые), при одновременном появлении у данного нейрона могут привести к его возбуждению при условии, что суммированный ВПСП достигает или превышает критический уровень деполяризации.
3. Явление окклюзии: один и тот же нейрон может передавать сигналы на ряд других нейронов, в результате чего возникает определенный эффект (например, происходит активация 10 мышечных волокон в мышце, вследствие чего мышца развивает напряжение, равное 100 мгс). Второй нейрон возбуждает тоже 10 других волокон (100 мгс). Но если оба нейрона возбуждать одновременно, то суммарная активность мышцы будет 180 мгс. Почему? Оказывается, часть волокон у них были общими (т. с. нейрон 1 и 2 передавали информацию на одни и те же волокна). Это явление получило название окклюзии или закупорки.
4. Трансформация ритма возбуждения. В отличие от скелетной мышцы или аксона нейрон способен Tpai формировать ритм возбуждений, приходящих к нему. Например, поступает импульс, идущий с частотой 25 Гц, а нейрон в ответ на это, возбуждаясь, генерирует 50 имп/сек (50 Гц), или наоборот, поступает 100 имп/сек, а выходят 40 имп/сек.
5. Последействие: один из вариантов этого свойства - длительное циркулирование импульсов но «нейронной ловушке». Итальянский физиолог Лоренто де Но обнаружил это явление: поступивший импульс может минутами или часами пробегать небольшой отрезок нейронной цепи. Благодаря этому, как полагают некоторые авторы, происходит перевод следа (энграммы) из краткосрочной памяти в долгосрочную.
6. Утомление нервных центров: это одно из важных свойств ЦНС. Оно обусловлено особенностями синаптической передачи в ЦНС: при длительном возбуждении одного и того же нейрона в синапсе может снизиться содержание медиатора, что приведет к снижению работоспособности нейрона.
(Торможение в центральной нервной системе.Виды торможения, его механизмы, значение)
Впервые идею о том, что в ЦНС помимо процессов возбуждения существует процесс торможения, высказал И.М. Сеченов.
Если рассмотреть «архитектуру» использования тормозных нейронов при организации нейронных сетей, цепей и рефлекторных дуг, то можно выделить ряд вариантов этой организации (это отражается в названии данного вида торможения).
1. Реципрокное торможение. Как пример: сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на альфа-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона разгибателя. Явление открыто Ч. Шеррингтоном.
2. Возвратное торможение. Альфа-мотонейрон, к примеру, посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС - она заканчивается на тормозном нейроне (клетка Реншоу) и активирует ее. Тормозной нейрон вызывает торможение альфа-мотонейрона, который запустил всю эту цепочку. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя тормозит.
3. Существует ряд вариантов возвратного торможения, в частности, латеральное торможение. Суть его сводится к тому, что активируется, например, фоторецептор, он активизирует биполярную клетку, и одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглиозной клетке. Этим самым происходит «вытормаживание» информации в соседних участках. Таким способом создаются условия для четкого видения предмета (две точки на сетчатке рассматриваются как две раздельные точки в том случае, если между ними есть невозбужденные участки).
(Общие принципы координационной деятельности ЦНС)
Координация - это объединение действий в единое целое, объединение различных нейронов в единый функциональный ансамбль, решающий конкретную задачу. Координация способствует реализации всех функций ЦНС. Выделяют следующие принципы координации (их много, в лекции даются наиболее важные).
1. Явление конвергенции (концентрации) или принцип общего конечного пути. Многие нейроны оказывают свое воздействие на один и тот же нейрон, т. с. имеет место схождение потоков импульсов к одному и тому же нейрону. Ч. Шеррингтон называл это «принцип общего конечного пути». Например, сокращение мышцы (за счет возбуждения альфа-мотонейрона) можно вызвать путем растяжения этой мышцы (рефлекс мышечных веретен) или путем раздражения кожных рецепторов (сгибательный рефлекс) и т.п.
2. Явление дивергенции (иррадиации). Каждый нейрон за счет вставочных нейронов и многочисленных ветвлений (дивергенции) дает поток импульсов ко многим нейронам. Если бы этот механизм не ограничивался торможением, то благодаря дивергенции отсутствовала бы возможность координации работы ЦНС. Но тормозные процессы ограничивают дивергенцию и делают процессы управления точными. Когда торможение снимается, то имеет место полнаядискоординация в деятельности ЦНС (например, при столбняке).
3. Принцип рсципрокной иннервации (см. Торможение; реципрокное торможение).
4. Принцип обратной связи и копий эфферентаций. Это один из важнейших принципов координации: невозможно точно координировать, управлять, если отсутствует обратная связь, т.е. данные о результатах управления. Осуществляется эта связь за счет потока импульсов с рецепторов. Этот принцип широко обсуждается в физиологии ЦНС, о нем уже говорил И.М. Сеченов, много внимания ему уделил П.К. Анохин.
Копия эфферентации. Для управления важно иметь информацию о том, какие команды посылаются на периферию. Известно, что в системах, управляющих скелетными мышцами, каждый отдел, посылая сигнал управления к работающей мышце, одновременно сообщает об этом вышележащему отделу. Это вариант обратной связи.
5. Принцип доминанты. Был открыт А.А. Ухтомским. Речь идет о том, что среди рефлекторных актов, которые могут быть выполнены в данный момент времени, имеются рефлексы, реализация которых представляет наибольший «интерес» для организма, т.е. они в данный момент времени самые важные. Поэтому эти рефлексы реализуются, а другие - менее важные - тормозятся.
6. Пластичность нервных центров. Для врача особенно важно знание этого свойства (принципа): при повреждении отдельных центров мозга их функция может перейти к другим структурам мозга (конечно, если повреждение центра не связано с наступлением смерти, что, например, бывает при нарушении дыхательного центра). Замещение утраченной функции - важнейшее приобретение ЦНС (известно, что нейроны ЦНС, как правило, не восстанавливаются) - оно позволяет восстанавливать утраченные свойства. Показано, что процесс возмещения утраченных функций осуществляется при обязательном участии коры больших полушарий: у животных, которым после восстановления нарушенных функций удаляли кору, вновь имела место утрата этой функции.
7. Принцип субординации или соподчинения. В ЦНС имеют место иерархические взаимоотношения - начальник (кора) и подчиненные (сверху вниз - базальные ганглии, средний мозг, продолговатый, спинной) и соподчинение - нижележащий отдел подчиняется указаниям вышележащего отдела.
Информационная функция нервной системы. Процесс восприятия сигнала, обработка его и посылка к исполнительной системе связаны с кодированием, «шифрованием» информации. Это происходит уже на уровне отдельной нервной клетки.
Для процесса передачи информации большое значение имеет скорость ее передачи.Важное значение в передаче информации имеет также надежность. Для обеспечения надежности передачи информации в нервной системе используется принцип структурной и функциональной избыточности. Структурная избыточность выражается в дублировании каналов связи, дублировании элементами, реагирующими на данный сигнал, дублировании системами, включающимися в реакцию. Функциональная избыточность обеспечивается «излишним» числом импульсов в разряде нервной клетки, существенным повышением возбудимости нервных структур и др.
Считают, что нервные импульсы передают информацию двоичным кодом (наличие импульса - отсутствие импульса). Из такого представления следует, что количество информации определяется числом импульсов в единицу времени.
(Сравнительная характеристика симпатического и парасимпатического отделов вегетативной нервной системы)
(симпатический)
Центральные структуры симпатического отдела вегетативной нервной системы расположены в спинном мозге.Они занимают пространство боковых рогов серого вещества от восьмого шейного сегмента до второго-третьего поясничного (спинномозговой центр Якобсона). Миелинизированные аксоны этого центра выходят в составе передних корешков спинного мозга.
Периферическая часть симпатического отдела состоит из двух пограничных стволов — цепочек паравертебральных ганглиев, лежащих по краям позвоночника. Ганглии в цепочке связаны между собой межузловыми ветвями (коннективами). Существуют и комиссуральные связи между симметричными ганглиями. В шейном и нижнем крестцовом отделах симпатического ствола преганглионарные нервы подходят к ганглиям не из своих сегментов спинного мозга, а из ниже- или вышележащих сегментов через коннективы ствола. В этом случае веточки проходят через ганглии, не переключаясь в них и оставаясь миелинизированными.
В шейном отделе имеются три шейных узла, образовавшиеся в онтогенезе при слиянии восьми симпатических ганглиев. Два верхних шейных узла иннервируют сонную артерию, глотку, пищевод, слюнные и щитовидные железы и сердце. Нижний шейный узел, в свою очередь, сливается с верхним грудным симпатическим узлом, образуя крупный звездчатый ганглий. Звездчатый ганглий иннервирует позвоночную артерию, органы грудной полости (пищевод, трахею, вилочковую железу, аорту) и сердечную мышцу.
В грудном отделе имеется 10—12 ганглиев. Постганглионарные ветви первых пяти из них направляются к плевре, сердечному, легочному и аортальному сплетениям. Узлы с 6-го по 9-й образуют большой чревный нерв, который, пройдя в брюшную полость, оканчивается в превертебральных узлах самого крупного нервного сплетения брюшной полости — чревного или солнечного сплетения. В его состав входят как симпатические, так и парасимпатические волокна. От чревного сплетения радиально отходит множество нервов, образующих вторичные сплетения (отсюда и название — солнечное сплетение). В узлах солнечного сплетения оканчиваются волокна многих внутренностных нервов. От узлов этого сплетения берут начало постганглионарные нервы, иннервирующие почти все органы брюшной полости. Последние 2—3 узла грудного отдела своими ветвями формируют малый чревный нерв, который также уходит к узлам солнечного сплетения.
В брюшной части пограничного симпатического ствола имеется 4—5 поясничных узла, от которых идут ветви к брюшной части аорты, половым органам, брыжеечному ганглию, кишечнику. В тазовой части симпатического ствола лежат 4 крестцовых узла и один непарный копчиковый узел. Через тазовое сплетение они иннервируют тазовые органы.
Симпатические афферентные (чувствительные) волокна от внутренних органов, несущие информацию от многочисленных интерорецепторов, идут в составе симпатических нервов и вступают в спинной мозг по его задним корешкам, как и чувствительные соматические афференты.
Окончания симпатических волокон выделяют в качестве медиатора норадренилин и адреналин.
Симпатический отдел вегетативной нервной системы увеличивает свою активность при необходимости мобилизации ресурсов организма. Под действием импульсов, приходящих по симпатическим нервам, увеличивается частота и сила сердечных сокращений, сужается просвет кровеносных сосудов, повышается кровяное давление, тормозится двигательная и секреторная активность пищеварительной системы.
Высшим центром, согласующим работу соматических и вегетативных функций, является кора больших полушарий. В ней имеются проекции как парасимпатических, так и симпатических нервов. Чувствительные пути вегетативных органов проецируются в лимбическую и ростральные части коры (орбитальная, двигательная зоны). Эти проекции строятся на топическом принципе — рядом расположенные органы проецируются в соседние зоны коры. Парасимпатические и симпатические проекции одних и тех же органов проецируются в одни и те же или близко расположенные участки коры. Однако парасимпатические проекции в коре представлены гораздо шире, чем симпатические.
Филогенез. Выделение вегетативной нервной системы из общей структуры ЦНС начинается уже у беспозвоночных. У кольчатых червей из клеток подглоточных узлов выделяются самостоятельные ганглии, связанные с кишечной трубкой. Разделение вегетативной нервной системы на симпатическую и парасимпатическую наблюдается уже у насекомых.
Низшие позвоночные (круглоротые) имеют блуждающий нерв и висцеральные (вегетативные) ветви спинномозговых нервов, однако цепочки симпатических ганглиев у них не наблюдается. Большая часть окончаний блуждающего нерва являются чувствительными. У низших рыб появляются сегментарные ганглии, связанные со спинномозговыми нервами и иннервирующие внутренние органы. Начиная с костистых рыб появляются продольные связи между ганглиями и образуется характерный для высших позвоночных парный пограничный симпатический ствол.
В различных группах позвоночных существуют свои особенности строения вегетативной нервной системы. У млекопитающих ярко проявляется тенденция к слиянию симпатических ганглиев. Клетки ганглиев периферических отделов вегетативной нервной системы возникают из ганглиозной пластинки. В процессе эмбрионального развития эти клетки перемещаются по ходу соответствующих нервов на периферию. В симпатическом отделе они перемещаются по ходу спинномозговых нервов, в парасимпатическом — по ходу блуждающего и тазового нервов. Покинув спинной мозг, симпатические нейроны располагаются двумя тяжами вдоль развивающегося позвоночника, образуя пограничные симпатические стволы. Некоторые из них мигрируют дальше, образуя превертебральные ганглии. Формирование периферических ганглиев значительно отстает от формирования структур центральной нервной системы.
Онтогенез. В онтогенезе парасимпатическая нервная система возникает раньше, чем симпатическая. Сначала наблюдается формирование чувствительных ганглиев и миграция нервных элементов на периферию по их нервам. Так, в сердце, на стадии зародыша 7, 5—11 мм, уже существует интрамуральное парасимпатическое сплетение, в то время как симпатические волокна прорастают к нему на стадии 17—23 мм.
На стадии 9-15 мм у зародыша наблюдается две пары параллельно идущих тяжей — закладки пограничных симпатических стволов, и лежащие вентральнее их закладки превертебральных сплетений брюшной полости. У зародыша 19 мм формируется шейное сплетение и симпатический ствол. С окончанием плодного периода симпатическая нервная система не заканчивает своего формирования. До трехлетнего возраста происходит интенсивный рост клеток, увеличение диаметра нервных волокон, увеличение размеров ганглиев.
(парасимпатический)
Центральные структуры парасимпатического отдела вегетативной нервной системы расположены в стволе мозга(средний мозг,
Варолиев мост и продолговатый мозг) и в крестцовом отделе спинного мозга.
Периферические части образованыэктрамуральными и интрамуральными ганглиями и нервами.
Из среднего мозга парасимпатические ветви уходят в составе глазодвигательного нерва (III пара). Затем преганглионарные волокна направляются к ресничному экстрамуральному ганглию глазницы. Постганглионарные волокна этого ганглия иннервируют гладкую мускулатуру ресничного тела и кольцевых мышц зрачка, т.е. являются двигательными.
Варолиев мост покидается парасимпатическими волокнами в составе лицевого нерва (VII пара). На периферии они образуют преганглионарные веточки нескольких экстрамуральных узлов, иннервирующих железы слизистой оболочки носа и нёба, слезные железы, подчелюстную и подъязычную слюнные железы. Таким образом, парасимпатические веточки лицевого нерва являются секреторными.
Из продолговатого мозга в составе языкоглоточного нерва (IX пара) уходят также секреторные парасимпатическое ветви, которые направляются к ушному экстрамуральному ганглию, иннервирующему околоушные слюнные железы и железы слизистой щек и губ.
Блуждающий нерв (X пара) является самой значительной частью парасимпатического отдела вегетативной нервной системы. Его ветви направляются к сердечному, бронхиальному и чревному сплетениям, а также к интрамуральным узлам в стенках внутренних органов грудной, брюшной полостей и полости большого таза.
Парасимпатические ветви крестцовой части спинного мозга берут начало в боковых рогах серого вещества второго—четвертого крестцовых сегментов и направляются к превертебральным ганглиям нижнего подчревного сплетения и интрамуральным ганглиям органов малого таза (см. рис. 19).
Окончаниями парасимпатического отдела вегетативной нервной системы выделяется медиатор ацетилхолин.
Парасимпатический отдел вегетативной нервной системы регулирует работу внутренних органов в условиях покоя. Его активация способствует снижению частоты и силы сердечных сокращений, снижению кровяного давления, увеличению как двигательной, так и секреторной активности пищеварительного тракта.
(Мышечный тонус, механизмы регуляции. Роль различных отделов центральной нервной системы в регуляции тонуса мышц)
Мышечный тонус - легкое напряжение мышц в отсутствии нагрузки, при котором она выглядит плотной.
Скелетные мышцы всегда находятся в состоянии некоторого напряжения. Постоянное незначительное напряжение мышц, не сопровождающееся признаками утомления, называется мышечным тонусом. Односторонняя перерезка у спинальной лягушки, подвешенной на крючке штатива, чувствительных (задних) корешков спинного мозга, в которых проходят афферентные нервные волокна, иннервирующие соответствующую заднюю лапку, приводит к исчезновению мышечного тонуса этой лапки и она распрямляется. К аналогичному эффекту приводит перерезка передних (двигательных) корешков или разрушение спинного мозга. Эти опыты свидетельствуют о том, что при разрушении основных звеньев рефлекторного кольца (афферентных и эфферентных путей, нервных центров) мышечный тонус исчезает. Следовательно, мышечный тонус имеетрефлекторную природу.
Источником возбуждений, поддерживающих мышечный тонус, являются проприорецепторы. В скелетных мышцах имеются три вида проприорецепторов:
• мышечные веретена, расположенные среди мышечных волокон;
• сухожильные рецепторы Гольджи, расположенные в сухожилиях;
• пачиниевы тельца, расположенные в фасциях, сухожилиях, связках.
Особое значение в регуляции мышечного тонуса имеют мышечные веретена и сухожильные рецепторы Гольджи.
Мышечные веретена представляют собой небольшие продолговатые образования, напоминающие своим внешним видом прядильные капсулы мышечного веретена находится пучок мышечных волокон, которые называются интрафузальными, т. к. они расположенными внутри веретена в отличие от обычных мышечных волокон, которые называются зкстрафузальными.
Каждое интрафузальное волокно состоит из трех частей:
• его центральная часть называется ядерной сумкой, в которой находятся ядра мышечной клетки;
• два периферических участка, которые имеют поперечную исчерченность и обладают способностью сокращаться;
• миотрубки, расположенные между ядерной сумкой и периферическими участками.
Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона-первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуя вторичные рецепторные окончания.
В мышце мышечное веретено одним концом прикрепляется к экстрафузальному мышечному волокну, а другим - к сухожилию этого волокна. Таким образом, мышечное веретено расположено в мышце параллельно экстрафузальным мышечным волокнам
При снижении тонуса экстрафузального волокна увеличивается его длина, что приводит к растяжению и раздражению первичных и вторичных рецепторных окончаний, для которых растяжение является адекватным раздражителем.
Возбуждение от рецепторных окончаний по афферентным волокнам поступает в спинной мозг к мотонейронам, расположенным в передних рогах. Мотонейроны спинного мозга принято подразделять на альфа- и гамма-мотонейроны (так как их аксоны относятся к А-альфа и А-гамма нервным волокнам). Возбуждение от альфа-мотонейронов поступает к экстрафузальным мышечным волокнам, вызывая их сокращение - тонус восстанавливается. Избыточное сокращение экстрафузальных мышечных волокон приводит к растяжению сухожильных рецепторов Гольджи, так как они прикрепляются к мышце последовательно. В них возникает возбуждение, которое поступает к тормозным вставочным нейронам спинного мозга, а от них к альфа-мотонейронам. Активность альфа-мотонейронов при этом снижается, уменьшается импульсация, идущая от них к экстрафузальным мышечным волокнам, тонус несколько снижается.
Высокая возбудимость мышечного веретена поддерживается за счет специального механизма, который образован сократительными элементами, расположенными в периферических участках интрафузальных волокон по обе стороны от ядерной сумки. Сокращение этих участков вызывает растяжение ядерной сумки и миотрубок, что приводит к возбуждению рецепторных окончаний и увеличению потока афферентных возбуждений к альфа-мотонейронам. Степень сокращения сократительных элементов интрафузальных волокон регулируется гамма-мотонейронами спинного мозга. Импульсы, приходящие по гамма-афферентным волокнам, вызывают сокращение периферических участков интрафузальных волокон.
Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальным или простейшим. Спинальный тонус характеризуется очень слабой выраженностью тонического напряжения. Такой тонус не может обеспечить поддержание позы животного и акт ходьбы, но он достаточен для осуществления простейших спинальных рефлексов.
Перерезка у животного (например, у кошки) ствола мозга между передними и задними буграми четверохолмия (операция перерезки ствола мозга называется децеребрацией) вызывает особое состояние скелетной мускулатуры, которое называетсядецеребрационной ригидностью или контрактильным тонусом. Это состояние характеризуется резким повышением тонуса разгибательной мускулатуры. Конечности такого животного сильно вытянуты, голова запрокинута, спина выгнута. Это состояние называется опистотонусом. Необходимо приложить большое усилие, чтобы согнуть у такого животного конечность в суставе.
Контрактильный тонус имеет рефлекторную природу. Это доказывается тем, что при перерезке передних или задних корешков спинного мозга, иннервирующих конечность, ригидность мускулатуры этой конечности исчезает.
Важную роль в возникновении контрактильного тонуса играет дорсальное вестибулярное ядро продолговатого мозга (ядро Дейтерса), которое возбуждается импульсами от рецепторов вестибулярного аппарата. При раздражении вестибулярного ядра ригидность усиливается, а при его разрушении ригидность уменьшается. Полагают, что это ядро оказывает влияние на ретикулярную формацию продолговатого мозга, которая оказывает неодинаковое влияние на нейроны спинного мозга. Раздражение медиальных отделов ретикулярной формации приводит к торможению рефлексов спинного мозга (тормозящая ретикуло-спинальная система), а раздражение латеральных отделов вызывает активацию нейронов спинного мозга (облегчающая ретикуло-спинальная система).
Ядро Дейтерса продолговатого мозга оказывает активирующее влияние на облегчающую ретикуло-спинальную систему и угнетает тормозную ретикуло-спинальную систему. Активность вестибулярного ядра тормозится красным ядром среднего мозга. При децеребрации происходит разобщение красного и вестибулярного ядер, в результате чего снижается тормозящее влияние красного ядра на вестибулярное ядро. В этих условиях оно оказывает сильное активирующее влияние на облегчающую ретикуло-спинальную систему. Поток импульсов от нее поступает в спинной мозг к гамма-мотонейронам, а от них к сократительным участкам интрафузальных волокон. Сокращение этих участков вызывает раздражение проприорецепторов. Поток возбуждений от них поступает к альфа-мотонейронам спинного мозга, а от них - к экстрафузальным мышечным волокнам, вызывая резкое увеличение их тонуса.
Разрушение красного ядра у интактного животного приводит к развитию гипертонуса мышц-разгибателей, а при его раздражении - к снижению тонуса.
Обнаружены и прямые связи красного ядра и ядра Дейтерса с мотонейронами спинного мозга. Показано, что ядро Дейтерса тормозит мотонейроны мышц-сгибателей и возбуждает мотонейроны мышц-разгибателей. Красное ядро оказывает тормозное влияние на мышцы-разгибатели и активирующее влияние на мышцы-сгибатели. Следовательно, при децеребрации, когда красное ядро отделено от ядра Дейтерса, создаются все условия для повышения тонуса мышц-разгибателей.
На ядро Дейтерса тормозное влияние оказывает и мозжечок. Удаление червячной зоны мозжечка у животного, находящегося в состоянии децеребрационной ригидности, вызывает большее растормаживание ядра Дейтерса и дальнейшее увеличение тонуса разгибателей. Электрическое раздражение червячной зоны, напротив, приводит к уменьшению тонуса этих мышц за счет активации тормозных влияний мозжечка на ядро Дейтерса.
При перерезке головного мозга выше промежуточного мозга у животного возникает особое изменение тонуса - мышцы становятся пластичными (воскоподобными), при этом конечностям можно легко придать любое положение, которое они могут сохранять длительное время. Такое состояние называется пластическим тонусом или восковой ригидностью. Пластический тонус имеет рефлекторное происхождение: после перерезки чувствительных нервов, иннервирующих конечность, все проявления пластического тонуса на этой конечности исчезают.
В возникновении пластического тонуса определенную роль играет черная субстанция среднего мозга. Черная субстанция функционально связана с базальными ганглиями - бледным шаром и полосатым телом. Нейроны черной субстанции синтезируют медиатор дофамин. Аксоны этих нейронов подходят к полосатому телу, которое также содержит дофамин. Повреждение черной субстанции, вызывающее дегенерацию дофаминергических путей к полосатому телу, сопровождаетеся заболеванием - болезнью Паркинсона. Одним из симптомов этой болезни является восковидная ригидность, которая обусловлена, по-видимому, гиперактивностью базальных ганглиев, возникающей при повреждении дофаминергического (вероятно, тормозного) пути, идущего от черной субстанции к полосатому телу.
Кроме того, черная субстанция, по-видимому, оказывает на скелетные мышцы трофическое влияние, подобно симпатической нервной системе. При перерезке выше промежуточного мозга высвобождаются структуры, которые оказывают на черную субстанцию тормозящее влияние. В таких условиях черная субстанция затормаживается и перестает оказывать достаточное трофическое влияние на скелетные мышцы и они становятся пластичными.
Пластический тонус может возникнуть не только при перерезке мозга, но и, например, при отравлении некоторыми ядами, при заболевании нервной системы, а также под влиянием гипноза. У человека пластический тонус может проявляться при особом состоянии нервной системы, которое называется каталепсией или восковой ригидностью. Человек в таком состоянии на некоторое время как бы цепенеет, причем, в неестественной позе и не меняет ее в течение долгого времени.
Большое значение в регуляции мышечного тонуса имеют базальные ядра - бледный шар и полосатое тело, которые образуют стриопаллидарную систему. Эти структуры регулируют активность всех нижележащих отделов ЦНС, участвующих в регуляции мышечного тонуса, обеспечивая адекватное перераспределение тонуса мышц при различных видах деятельности. При поражении экстрапирамидной системы, составной частью которой являются базальные ядра, возникают нарушения регуляций тонуса мускулатуры, что приводит к развитию так называемых дрожательных параличей (паркинсонизму, атетозу, хорее и др.).
Главную роль в приспособительной регуляции мышечного тонуса выполняет кора головного мозга. С ее участием и участием других структур, регулирующих мышечный тонус, формируется нормальный или корковый тонус.
Регуляция тонуса скелетной мускулатуры осуществляется экстрапирамидной системой головного мозга. Быстрые (фазные) движения обеспечиваются активностью пирамидной системы.
В условиях целого организма пирамидная и экстрапирамидная системы включаются как единое целое в функциональную архитектуру приспособительных актов, обеспечивая необходимые фазные движения и адекватные изменения мышечного тонуса. Специфическое изменение тонуса мышц возникает при осуществлении статических и стато-кинетических рефлексов, направленных на поддержание позы.
Статические рефлексы делятся на рефлексы положения и рефлексы выпрямления. Рефлексы положения обеспечивают формирование тонуса мышц, необходимого для поддержания естественного положения тела в пространстве в состоянии покоя. Рефлексы выпрямления определяют перераспределение тонуса мышц, приводящее к восстановлению естественной для данного вида животного позы в случае ее изменения.
Стато-кинетические рефлексы направлены на сохранение позы в пространстве при ускорениях прямолинейного и вращательного характера. Эти рефлексы проявляются при вращении, перемещении тела в горизонтальной и вертикальной плоскостях. Они возникают, в основном, в результате возбуждения рецепторов вестибулярного аппарата. Так, при вращательном движении наблюдается нистагм головы: вначале голова медленно поворачивается в сторону, противоположную направлению вращения, а затем быстро возвращается в. исходное положение. Такое же движения глазных яблок при вращении называется нистагмом глаз.
Перераспределение тонуса мышц шеи, туловища и конечностей происходит при быстром подъеме и спуске. Быстрый подъем сопровождается повышением тонуса сгибателей, а быстрый спуск - разгибателей конечностей. Эти рефлекторные реакции легко наблюдать при перемещении в скоростном лифте, поэтому они и называются лифтными рефлексами.
(Корковый отдел анализаторов, особенности его структуры и функции)
(Свойства)
1. Каждая сенсорная система ( каждый анализатор) имеет проекцию в кору больших полушарий. Корковый отдел анализаторов имеет центральную часть и окружающую ее ассоциативную зону (по представлению И. П. Павлова — “ ядро” и рассеянные элементы). Центральная часть коркового отдела анализатора состоит из высокодифференцированных в функциональном отношении нейронов, которые осуществляют высший анализ и синтез информации, поступающей к ним. Ассоциативные корковые зоны представлены менее дифференцированными нейронами, способных к выполнению простейших функций. Синтез и анализ афферентных импульсов этими клетками осуществляется в элементарной, примитивной форме.
2. Одной из общих черт организации сенсорных систем является принцип двойственной проекции их в кору больших полушарий. Этот принцип тесно связан с многоканальностью проводящих путей и выражается в осуществлении двух различных типов корковых проекций, которые можно разделить на первичные и вторичные проекции. Первичные и вторичные проекционные зоны окружены ассоциативными корковыми зонами той же сенсорной системы. Примером двойственной проекции в коре головного мозга может служить представительство вкусового анализатора. Его первичная корковая проекция представлена, по- видимому, орбитальной областью коры, так как именно здесь при раздражении рецепторов языка вызванные ответы возникают с самым коротким латентным периодом и имеют самую высокую амплитуду. Вторичной проекционной областью коры вкусового анализатора является соматосенсорная область. Здесь вызванные ответы возникают значительно позже, чем в орбитальной области, и амплитуда их меньше.
3. Взаимодействие анализаторов на корковом уровне осуществляется за счет ассоциативных корковых зон и за счет наличия полимодальных нейронов.
Взаимодействие анализаторов. Деятельность одних анализаторов находится в зависимости от деятельности других, причем, может наблюдаться как усиление деятельности анализатора, так и ее ослабление.
Взаимодействие анализаторов осуществляется на различных уровнях — спинальном, ретикулярном и таламо- кортикальном. Особенно широкая интеграция сигналов наблюдается в нейронах ретикулярной формации. Интеграция сигналов высшего порядка осуществляется на корковом уровне. В результате множественных связей с нижележащими уровнями анализаторов и неспецифических систем многие нейроны коры приобретают способность отвечать на сложные комбинации сигналов различной природы. Это особенно свойственно клеткам ассоциативных областей, а также моторной зоне коры больших полушарий. Пирамидные клетки этой области коры являются общим конечным путем слуховых, зрительных, тактильных и других сигналов.
(Зрительный анализатор. Структурно-функциональные особенности рецепторного аппарата. Восприятие цветов.)
Периферический отдел зрительного анализатора - фоторецепторы, расположенные на сетчатой оболочке глаза. Нервные импульсы по зрительному нерву (проводниковый отдел) поступают в затылочную область — мозговой отдел анализатора. В нейронах затылочной области коры большого мозга возникают многообразные и различные зрительные ощущения.
Глаз состоит из глазного яблока и вспомогательного аппарата. Стенку глазного яблока образуют три оболочки: роговица, склера, или белочная, и сосудистая. Внутренняя (сосудистая) оболочка состоит из сетчатки, на которой расположены фоторецепторы (палочки и колбочки), и ее кровеносных сосудов.
В состав глаза входят рецепторный аппарат, находящийся в сетчатке, и оптическая система. Оптическая система глаза представлена передней и задней поверхностью роговой оболочки, хрусталиком и стекловидным телом. Для ясного видения предмета необходимо, чтобы лучи от всех его точек падали на сетчатку.Приспособление глаза к ясному видению разноудаленных предметов называют аккомодацией. Аккомодация осуществляется путем изменения кривизны хрусталика. Рефракция – преломление света в оптических средах глаза.
Существуют две главные аномалии преломления лучей в глазу: дальнозоркость и близорукость.
Поле зрения — угловое пространство, видимое глазом при фиксированном взгляде и неподвижной голове.
На сетчатке расположены фоторецепторы: палочки (с пигментом родопсин) и колбочки (с пигментом йодопсин). Колбочки обеспечивают дневное зрение и восприятие цвета, палочки – сумеречное, ночное зрение.
Человек обладает способностью различать большое количество цветов. Механизм цветовосприятия по общепринятой, но уже устаревшей трехкомпонентной теории заключается в том, что в зрительной системе имеются три датчика, чувствительных к трем основным цветам: красному, желтому и синему. Поэтому нормальное цветовосприятие называется трихромазией. При определенном смешении трех основных цветов возникает ощущение белого цвета. При нарушении работы одного или двух датчиков основных цветов правильного смешения цветов не наблюдается и возникают нарушения цветовосприятия.
Различают врожденную и приобретенную формы цветоаномалии. При врожденной цветоаномалии чаще наблюдается снижение чувствительности к синему цвету, а при приобретенной — к зеленому. Цветоаномалия Дальтона (дальтонизм) заключается в снижении чувствительности к оттенкам красного и зеленого цветов. Этим заболеванием страдают около 10 % мужчин и 0,5 % женщин.
Процесс восприятия цвета не ограничивается реакцией сетчатки, а существенно зависит от обработки полученных сигналов мозгом.
(Рефракция глаза, ее аномалии. Физиологические механизмы аккомодации глаза)
Рефракция (физическая рефракция) — преломляющая сила оптической системы глаза, которая измеряется условной единицей — диоптрией. За одну диоптрию принята преломляющая сила стекла с главным фокусным расстоянием в 1 м. Диоптрия — величина, обратная главному фокусному расстоянию. Средняя преломляющая сила нормального глаза может варьировать в пределах от 52,0 до 68,0 D. В офтальмологии важна не рефракция оптической системы глаза, а ее способность фокусировать лучи на сетчатке. Поэтому используется понятие клиническая рефракция, т.е. положение заднего главного фокуса оптической системы глаза по отношению к сетчатке.
(аномалии)
У большинства людей глаз имеет шарообразную форму, максимально приспособленную для того, чтобы световые лучи фокусировались на сетчатке при любой преломляющей силе хрусталика, изменяющейся в процессе аккомодации. При нормальной рефракции на сетчатке одинаково четко проецируются изображения далеких и близких предметов, а проявление такой универсальной рефракционной способности определяют термином эмметропия. Аномалии рефракциивозникают при изменениях формы глаза, проявляющихся в удлинении его оптической оси (близорукость, или миопия) или в ее укорочении (дальнозоркость, илигиперметропия). При удлиненной форме глаза, характерной для миопии, обычная преломляющая способность его оптической системы оказывается избыточной, в связи с чем изображение фокусируется перед сетчаткой и субъективно воспринимается нечетким. Дефект зрительного восприятия ощущается только при рассматривании удаленных предметов, тогда как световые лучи, отраженные от близко находящихся предметов, фокусируются точно на сетчатке благодаря ограничению механизма аккомодации. Например, при чтении близоруким людям нужна меньшая, чем людям с нормальной рефракцией, преломляющая сила хрусталика, поэтому нагрузка на ресничные мышцы у них снижается. Но для ясного видения далеких предметов близорукому человеку приходится носить очки с двояковогнутыми рассеивающими линзами, уменьшающими преломление световых лучей. При характерной для гиперметропии укороченной оптической оси глаза световые лучи фокусируются за сетчаткой, что принуждает ресничные мышцы к сокращению даже при рассмотрении далеких предметов. Для компенсации этой аномалии рефракции необходимы очки с двояковыпуклыми линзами, преломляющими световые лучи так, чтобы они фокусировались на сетчатке.
Наибольшей преломляющей способностью в оптической системе глаза обладает роговица (около 48,8 диоптрий), которая не участвует в аккомодации. Поверхность роговицы не идеально сферична почти у всех людей, ее кривизна по вертикали выражена больше, чем по горизонтали. При значительном отклонении формы роговицы от идеальной пропорции возникает аномалия рефракции, которая определяется как астигматизм. В результате такой аномалии световые лучи, отраженные от наблюдаемых объектов, преломляются не одинаково, поэтому часть лучей фокусируется на сетчатке, а остальные — за ее пределами, что ведет к искажению зрительного восприятия. Для исправления этого дефекта применяются индивидуально подобранные линзы очков с различной кривизной по вертикальному и горизонтальному меридианам.
(аккомодация)
Аккомодация — изменение преломляющей силы глаза или способность глаза фокусировать на сетчатке световые лучи, отраженные от рассматриваемых предметов, вне зависимости от расстояния между глазом и этими предметами, т.е. видеть хорошо и вдаль, и вблизи. Аккомодация приспосабливает глаз четко различать предметы, располагающиеся между ближайшей и дальнейшей точками ясного видения. То минимальное расстояние от глаза, на котором глаз еще может отчетливо различать предмет, принято называть ближайшей точкой ясного видения. Дальнейшая точка определяется наибольшим расстоянием, на котором ясно различим предмет при отсутствии аккомодации. Механизм аккомодации. Решающая роль принадлежит хрусталику и цилиарной мышце. Во время покоя цилиарной мышцы зонулярные волокна натянуты. Она связана, с одной стороны, с цилиарным телом, а с другой — с сумкой хрусталика и оказывает на последнюю некоторое давление, не позволяя ему принять более выпуклую форму. При сокращении аккомодационной мышцы расслабляются зонулярные волокна, уменьшаются силы, натягивающие капсулу хрусталика, и вследствие эластичности своих волокон он становится более выпуклым. Преломляющая сила глаза увеличивается, и изображение близкого предмета на сетчатке становится четким. Абсолютная и относительная аккомодация. Аккомодация, определяемая для одного глаза, называется абсолютной. Если зрение осуществляется двумя глазами (бинокулярно), то акт аккомодации обязательно сопровождается конвергенцией, т.е. сведением зрительных осей глаз на фиксируемом предмете (относительная аккомодация). Аккомодация и конвергенция у человека, имеющего эмметропию, обычно совершается параллельно и согласованно. Объем относительной аккомодации всегда несколько меньше абсолютного ее объема. Если запас аккомодации мал, то во время работы быстро возникает явление усталости — зрительное утомление. Причины нарушения аккомодационной способности. С возрастом аккомодационная способность глаза ослабевает. Это может быть обусловлено изменением физико-химического состава хрусталика, обеднением его водой, уплотнением, особенно в области ядра, потерей эластичности. Вследствие этого от глаза постепенно отдаляется ближайшая точка ясного видения. После 40 лет эта точка находится уже на довольно большом расстоянии, и поэтому для рассматривания мелких предметов их приходится не приближать, а отодвигать от глаза все дальше и дальше. Возникает пресбиопия, т.е. старческая дальнозоркость. Лицам с пресбиопией для зрительной работы на близком расстоянии необходима очковая коррекция.