

Обмен веществ и энергии в нервной ткани
Энергетический обмен нервной ткани
Для мозга характерна высокая интенсивность энергетического обмена с преобладанием аэробных процессов. При массе 1400 г (2% массы тела), он получает около 20% крови, выбрасываемой сердцем и приблизительно 30% всего кислорода, находящегося в артериальной крови. В отличие от других органов, головной мозг практически не располагает запасами кислорода. Резервный кислород мозга расходуется в течение 10-12 секунд, что объясняет высокую чувствительность нервной системы к гипоксии.
Максимальная скорость дыхания обнаружена в коре больших полушарий, минимальная – в спинном мозге и периферических нервах. Нейронам свойственен аэробный обмен, тогда как метаболизм нейроглии адаптирован и к анаэробным условиям. Интенсивность дыхания серого вещества в 4 раза выше, чем белого.
Максимальный энергетический обмен в мозге наблюдается к периоду окончания миелинизации и завершения процессов дифференцировки у детей в возрасте 4 лет, быстро растущая нервная ткань потребляет около 50% всего кислорода поступающего в организм.
Основным энергетическим субстратом нервной ткани является глюкоза, окисления которой обеспечивается ее энергией на 85-90%. Нервная ткань потребляет до 70% свободной глюкозы артериальной крови. В физиологических условиях 85-90% глюкозы метаболизирует аэробным путем, а 10-15% - анаэробным.
В качестве дополнительных энергетических субстратов нейроны и глиальные клетки могут использовать аминокислоты, в первую очередь глутамат и аспартат. В ранний постнатальный период в мозге также окисляются свободные жирные кислоты и кетоновые тела.
В экстремальных состояниях нервная ткань переключается на кетоновые тела (до 50% всей энергии).
Полученная энергия тратится в первую очередь:
на создание мембранного потенциала, который используется для проведения нервных импульсов и активного транспорта;
для работы цитоскелета, обеспечивающего аксональный транспорт, выделение нейромедиаторов, пространственной ориентации структурных единиц нейрона;
для синтеза новых веществ, в первую очередь нейромедиаторов, нейропептидов, а также нуклеиновых кислот, белков, липидов;
для обезвреживания аммиака.
Обмен углеводов нервной ткани
Нервная ткань инсулиннезависима, с высокой активностью гексокиназы (имеет низкую константу Михаэлиса Ментона) и низкой концентрацией глюкозы, глюкоза поступает из крови в нервную ткань постоянно, даже если в крови мало глюкозы и отсутствует инсулин. Основное направление – энергетический обмен.
Активность ПФШ нервной тканинижепо сравнению с печенью.. НАДФН2 используется при синтезе нейромедиаторов, аминокислот, липидов, гликолипидов, компонентов нуклеиновых кислот и для работы антиоксидантной системы.
Высокая активность ПФШ наблюдается у детей в период миелинизации и при травмах головного мозга.
Обмен белков и аминокислот нервной ткани
Нервная ткань характеризуется высоким обменом аминокислот и белков.
Скорость синтеза и распада белков в разных отделах головного мозга неодинакова. Белки серого вещества больших полушарий и белки мозжечка отличаются высокой скоростью обновления, что связано с синтезом медиаторов, БАВ, специфических белков. Белое вещество, богатое проводниковыми структурам, обновляется особенно медленно.
Аминокислоты в нервной ткани используется как:
источники синтеза белков, пептидов, некоторых липидов, ряда гормонов, биогенных аминов. В сером веществе преобладает синтез БАВ, в белом – белков миелиновой оболочки.
нейротрансмиттеры и нейромодуляторы. Аминокислоты и их производные участвуют в синаптической передаче (глу), в осуществлении межнейрональных связей.
Источник энергии. Нервная ткань использует в ЦТК аминокислоты глутаминовой группы и с разветвленной боковой цепью (лейцин, изолейцин, валин).
Для выведения азота. При возбуждении нервной системы за счет дезаминирования АМФ возрастает образование аммиака, который связывается с глутаминовой кислотой с образованием глутамина. Реакцию с затратой АТФ катализирует глутаминсинтетаза.
Аминокислоты глутаминовой группы имеют самый активный метаболизм в нервной ткани.
N-ацетиласпарагиновая кислота (АцА) является частью внутриклеточного пула анионов и резервуаром ацетильных групп. Ацетильные группы экзогенной АцА служат источником углерода для синтеза жирных кислот в развивающемся мозге.
Ароматические аминокислоты имеют особое значение как предшественники катехоламинов и серотонина.
Серосодержащие аминокислоты
Метионин является источником метильных групп и на 80% используется для синтеза белка.
Цистатионин важен для синтеза сульфитидов и сульфатированных мукополисахаридов.
Обмен аминокислот нервной ткани
Непосредственным источником аммиака в головном мозге служит трансаминирование и прямое дезаминирование аминокислот с участием глутаматдегидрогеназы и дезаминирование с участием АМФ–ИМФ цикла.
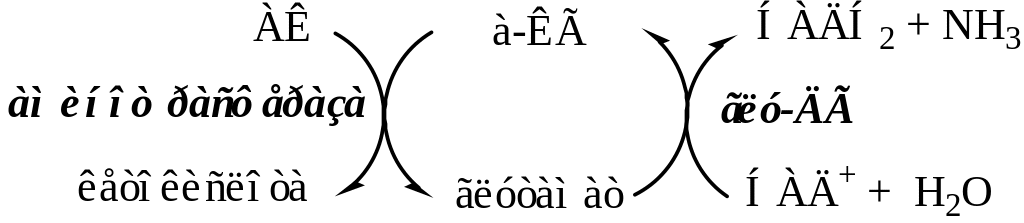

Обезвреживание токсичного аммиака в нервной ткани происходит с участием α-кетоглутарата и глутамата.
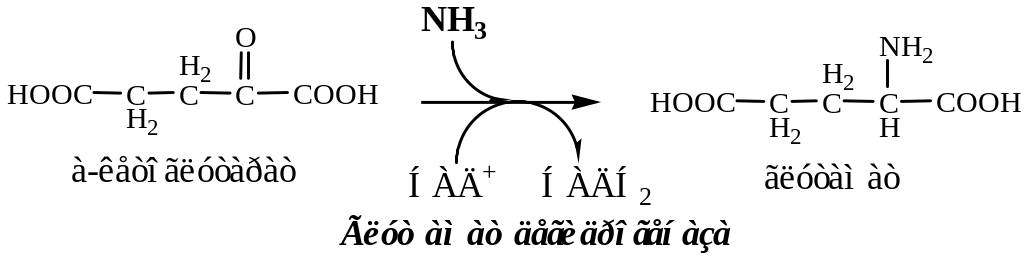

Липидный обмен нервной ткани
Особенность обмена липидов - не используются в качестве энергетического материала, а в основном идут на образование специфических липидов нервной ткани...
В нейронах серого вещества из всех фосфоглицеридов наиболее интенсивно обновляются фосфатидилхолины, и особенно фосфотидилинозитол, который является предшественником внутриклеточного посредника ИТФ, участвующего во внутриклеточной регуляции.
Обмен липидов в миелиновых оболочках протекает медленно, очень медленно обновляются холестерин, цереброзиды и сфингомиелины. У новорожденных холестерин синтезируется в самой нервной ткани, у взрослых этот синтез резко снижается, вплоть до полного прекращения.
ОБМЕН НЕЙРОМЕДИАТОРОВ