

5. Специфический иммунитет. Гуморальный ответ (выработка антител в-лимфоцитами). Патология иммуноглобулинов, связанная с нарушением их структуры, низким или высоким содержанием.
Гуморальный ответ связан с выработкой антител (иммуноглобулинов) — особых белков, циркулирующих в крови и других жидкостях организма и способных специфически связываться с чужеродными молекулами. Связывание с антителами инактивирует вирусы и бактериальные токсины, подавляя их способность связываться с рецепторами клеток-мишеней и проявлять свое инфицирующее и токсическое действие.
В-лимфоциты вырабатывают антитела — главные молекулы гуморального иммунитета. Развитие В-лимфоцитов происходит в костном мозге взрослых или в печени у плода.
Этот процесс не зависит от антигена, однако дифференцировка лимфоцитов во вторичных лимфоидных органах тесно связана с наличием антигена, под влиянием которого В-клетки синтезируют антитела, блокирующие данный антиген.
На мембранах зрелой В-клетки начинают образовываться рецепторы для антигена, причем этот процесс временный; он заканчивается образованием виргильной В-клетки, имеющей на мембранах полный набор структур, необходимых для взаимодействия с антигеном и хелперной Т-клеткой.
Клетки иммунологической памяти. Для иммунной системы характерно наличие памяти. Введение антигена в организм вызывает первичный иммунный ответ. Если тот же антиген ввести повторно то формируется вторичный иммунный ответ, причем реакция будет более сильной и продолжительной. Это обусловлено наличием в организме долго живущих клеток иммунологической памяти по отношению к данному антигену. В иммунизированном организме имеются Т- и В-клетки памяти, которые сами не дают ответа, но легко превращаются в активные клетки под действием соответствующего антигена. Т-клетки иммунологической памяти способны реагировать на более низкие дозы антигена, что повышает эффективность иммунного ответа.
Выделяют ряд патологических состояний связанных с нарушение структуры Ig, низким или высоким его содержанием. Например:
1. Нарушение конечного этапа дифференцировки В-клеток приводит к резкому снижению количества IgА, а в 20% случаев и IgG;
2. Гиперпродукция IgМ вызванная генетическим нарушением, при котором в В-клетках не происходит переключение с синтеза Ig М на синтез IgG, IgА, IgЕ. В нормальных условиях такое переключение обусловлено двумя факторами: связыванием ИЛ-4 с рецептором к ИЛ-4 на поверхности В-клеток и взаимодействием молекул СD40 на поверхности В-клеток с лигандом, СD40L, на активированных Т-клетках. Нарушение обусловлено мутацией в гене лиганда.
6. Клеточный ответ, обусловленный т-лимфоцитами. Т-клеточные антигенраспознающие рецепторы, белки главного комплекса гистосовместимости.
Клеточный ответ связан со специфическим узнаванием клетками иммунной системы антигенов, расположенных на поверхности других клеток организма (фрагменты вирусных белков и белков, образующихся при трансформации клеток).
Т-лимфоциты развиваются в тимусе и отвечают за клеточный иммунитет. Различают три основных вида Т-лимфоцитов:
Т-киллеры, или цитотоксические Т-лимфоциты, уничтожают клетки организма, зараженные вирусами, и вместе с В-лимфоцитами непосредственно участвуют в защите организма. Поэтому Т-киллеры и В-лимфоциты называют эффекторными клетками.
Т-хелперы и Т-супрессоры играют регуляторную роль, соответственно усиливая или подавляя размножение и созревание других клеток иммунной системы. Их объединяют в группу регуляторных клеток.
Т-лимфоциты могут узнавать инфицированную клетку, если соответствующий антиген расположен на ее поверхности. Взаимодействие с антигеном возможно только в комплексе с поверхностными маркерами – группой белков комплекса гистосовместимости МНС (от англ. Major histocompatibility complex).
Антиген, поглощенный антигенпрезентирующими клетками (АПК), деградирует на отдельные антигенные полипептиды, которые ассоциируются с белками МНС и перемещаются на поверхность клетки. К АПК, в частности, относятся дендритные клетки, макрофаги, а также В-клетки.
АПК представляют Т-хелперам фрагменты антигена (эпитопы), способные взаимодействовать с рецепторами Т-клеток.
Когда хелперные Т-клетки узнают «свой» антиген, они активируются и начинают продуцировать цитокины – белковые регуляторы иммунного ответа, осуществляющие межклеточные взаимодействия при иммунном ответе. Цитокины – небольшие белки или полипептиды. Они могут действовать как на клетку, которая их продуцирует, так и на другие вблизи расположенные клетки. К цитокинам относятся интерлейкины (ИЛ), интерфероны, факторы некроза опухолей, факторы роста, хемотоксические факторы и др.
Действие Тц-клеток на клетки-мишени. Главная функция Тц-клеток связана с уничтожением клеток, зараженных вирусами. При помощи специфического рецептора Тц-клетки распознают экзогенный антиген и присоединяются к клетке-мишени.
Для уничтожения инфицированных клеток Тц и НК имеют несколько механизмов воздействия. Одним из таких механизмов является воздействие на мембрану клетки-мишени гранул Тц или НК, содержащих белок перфорин и гранзимы — ферменты, ассоциированные с гранулами. Перфорин — мономерный белок, при воздействии на мембраны вызывает образование пор. В присутствии Са2+ мономеры перфорина связываются с мембраной клетки-мишени, полимеризуются и образуют трансмембранный канал. Это может привести к осмотическому шоку, неконтролируемому удалению внутриклеточных структур и, как следствие, гибели клетки. Гранзимы представляют собой набор цистеиновых протеиназ, расщепляющих полипептидные цепи по остатку аспарагиновой кислоты и вызывающих активацию ядерных протеиназ-каспаз. Те, в свою очередь, в результате ограниченного протеолиза активируют ядерную ДНК-азу и индуцируют программированную гибель клеток — апоптоз.
Т-супрессорные (Тс)-клетки. Некоторые Т-клетки способны подавлять те или иные механизмы иммунного ответа, поэтому их называют супрессорными клетками. Они образуют гетерогенную популяцию со смешанными функциями. Одна из субпопуляций Тс синтезирует и секретирует цитокин, способный ингибировать фактор роста. Клетки этой субпопуляции являются истинными супрессорами. Другие Тс-клетки совместно с Т-хелперами контролируют активность В-клеток и Тц-клеток — главных эффекторных клеток иммунной защиты, т.е. выполняют чисто регуляторную функцию без подавления клеточного роста.
Семейство Т-клеточных антигенраспознающих рецепторов.
Антитела связывают антигены в жидкостях организма (гуморальный иммунитет), а Т-лимфоциты взаимодействуют с антигенами на поверхности зараженных вирусами и измененных в результате опухолевой трансформации собственных клеток организма.
Т-лимфоциты узнают антигены только в комплексе с молекулами МНС I или II. На поверхности Т-лимфоцитов присутствуют рецепторы, распознающие антигены чужеродной клетки. Они представляют собой гликозилированные гетеродимеры, состоящие из двух неравнозначных цепей (α и β). Каждая цепь содержит два домена, один из которых имеет константную, а другой — вариабельную структуру. С-концевые участки каждой цепи встроены в плазматическую мембрану. Единственный антигенсвязывающий участок распологается между двумя вариабельными доменами. По своему строению рецепторы Т-клеток имеют много общего с иммуноглобулинами. Кроме того, у всех активированных Т-клеток рецептор нековалентно связан с белком ТЗ, состоящим из трех полипептидных цепей (γ, δ и ε). Этот белок участвует в передаче сигнала от антигенрецепторного комплекса внутрь клетки.
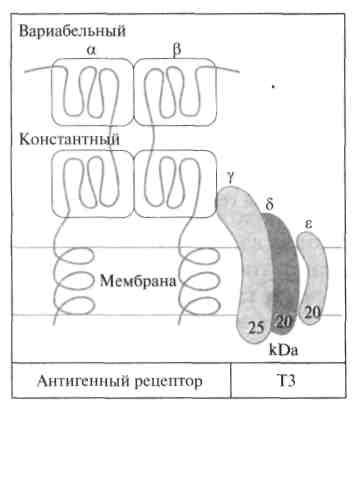
Рис.Строение Т-клеточного рецептора.