

Билет № 12.
4. Первая личиночная стадия возбудителя описторхоза: мирацидия.
5. (См. Билет №1.6)
Облегченная диффузия — специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом, или молекулой и переносят их через мембрану.
Активный транспорт. Этот механизм сопряжен с затратами энергии и служит для переноса веществ против их градиента концентрации. Он осуществляется специальными белками-переносчиками, образующими, так называемые, ионные насосы. Наиболее изученным является Nа+/К+-насос в клетках животных, активно выкачивающий ионы Nа наружу, поглощая при этом ионы К+ .
В сочетании с активным транспортом ионов в клетку через цитоплазматическую мембрану проникают различные сахара, нуклеотиды, аминокислоты.
Макромолекулы, такие, как белки, нуклеиновые кислоты, полисахариды, липопротеидные комплексы и др. сквозь клеточные мембраны не проходят, в отличие от ионов и мономеров.
6. Стартовым кодоном у эукариотических организмов является триплет AUG в мРНК, кодирующий метионин, с которого начинается образование полипептидной цепи в процессе трансляции.
*из 64 кодонов включения аминокислот в синтезирующуюся полипептидную цепь шифрует 61 триплет, а 3 остальных UAA, UAG, UGA не кодируют включение в белок аминокислот и первоначально были названы бессмысленными, или нонсенкодоном. Однако в дальнейшем было показано, что эти триплеты сигнализируют о завершении трансляции, и поэтому их стали называть терминируюшими, или стоп-кодонами.
**Синтез белка в большинстве случаев начинается с AUG-кодона, кодирующего метионин. Этот кодон обычно называют стартовым или инициаторным. Инициация трансляции предусматривает узнавание рибосомой этого кодона и привлечение инициаторной аминоацил-тРНК.
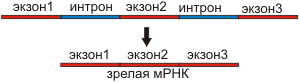
7. Многие гены состоят из экзонов (кодирующие участки) и интронов (некодирующие участки). При транскрипции с гена считывается РНК, несущая как экзоны, так и интроны. В процессе сплайсинга интроны вырезаются, а экзоны, сшиваясь, образуют зрелую РНК. (см. Бил. №10, 7, №11. 7) |
|
8. Что происходит с клетками на стадии созревания при оогенезе?
Созревание ооцита — это процесс последовательного прохождения двух делений мейоза. Как уже говорилось выше, при подготовке к первому делению созревания ооцит длительное время находится на стадии профазы I мейоза, когда и происходит его рост. Выход из профазы I мейоза приурочены к достижению самкой половозрелости и определяются половыми гормонами.
Из двух делений созревания первое у большинства видов является редукционным, так как именно в ходе этого деления гомологичные хромосомы расходятся по разным клеткам. Таким образом, каждая из разделившихся клеток приобретает половинный набор хромосом, где каждый ген представлен лишь одной аллелью.
Поскольку первому делению созревания предшествовала S-фаза, каждая из разошедшихся хромосом содержит двойное количество ДНК. Эти генетически идентичные хроматиды и расходятся по сестринским клеткам во втором делении созревания, которое является эквационным. После двух делений созревания число хромосом в каждой из клеток оказывается гаплоидным, а общее количество хроматина в каждом клеточном ядре будет соответствовать 1с.
9.Заражение эхинококкозом: проглатывая яйца, которые попадают на руки с шерсти собак или овец.
10. Половая структура популяции. Численное соотношение полов, т.е. половой состав, и особенно доля размножающихся самок в популяции, имеет большое значение для дальнейшего роста ее численности. Соотношение полов зависит, прежде всего, от биологии вида и сильно различается у моногамных (самец за сезон спаривается с одной самкой) и полигамных животных. Для первых (например, журавли, лебеди) нормой является соотношение полов 1:1. Для вторых (например, морские котики, павианы) типично преобладание самок. Среди моногамных животных почти постоянно имеются «резервные» самцы. Это уже половозрелые, но еще не размножающиеся животные; они представляют собой репродуктивный резерв популяции.
11. Строение оперона. Оперон - группа тесно связанных между собой генов, которые регулируют образование ферментов в организме. В состав оперона входит один или несколько структурных генов, которые определяют природу образующихся ферментов, и ген-оператор, который контролирует деятельность структурных генов и сам, в свою очередь, контролируется регуляторным геном, не входящим в состав оперона.
*Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать экспрессию (транскрипцию) этих генов. Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов). Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор. Все структурные гены, объединенные в оперон, имеют один операторный участок, локализованный на краю оперона, и координированно регулируются одним репрессором. Оперон представляет собой весьма рациональную и эффективную систему регуляции метаболического пути.
12. Популяционные волны, как эволюционный фактор.
Популяционные волны - это колебания численности особей в природных популяциях (это вспышки численности, периодические или непериодические значительные изменения числа особей в популяции). Масштабы колебаний численности у популяций разных видов обычно разные.
Они свойственны всем видам животных, растений и микроорганизмов. Популяционные волны могут быть сезонными (периодическими), генетически обусловленными и несезонными (апериодическими), обусловленными воздействием на популяцию непосредственно различных биотических и абиотических факторов. Так, увеличение кормовых ресурсов - растительности приводит к увеличению численности грызунов (например, леммингов), что, в свою очередь, через некоторое время приводит к увеличению численности хищников (лисиц, песцов). Последнее ведёт к истреблению жертвы (грызунов) и уменьшению кормовых ресурсов для хищников и в конечном итоге к уменьшению численности хищников. В XIX - XX веках отдельные «вспышки» численности организмов отмечались в популяциях кроликов в Австралии, домовых воробьев в Северной Америке, саранчи в Азиатско-Африканском регионе и т.п. После холодной зимы численность кроликов на одном из островов вблизи побережья Англии уменьшилась в 100 раз. Поэтому волны жизни опасны для выживания малочисленных популяций. Эволюционное значение популяционных волн сводится к случайным изменениям концентрации и частоты различных аллелей и генотипов, содержащихся в популяции. Изменение генофонда популяции происходит:
а) на подъёме популяционной волны (при возрастании численности популяции, усиливающиеся межпопуляционные масти зайцев и хищников) приводят к изменению генофонда популяции; б) на гребне популяционной волны (при наибольшей численности популяции): с возрастанием конкуренции возможно выселение особей за пределы ареала вида, где они испытывают действие новых (нетипичных) условий, которые могут существенно повлиять на частоту определённых аллелей.
*Важной причиной популяционных волн является также постепенное истощение пищевых ресурсов в связи с ростом популяции и последующее их восстановление после снижения численности популяции. Причинами резких непериодических снижений численности популяции могут также быть стихийные бедствия: засухи, пожары, наводнения. Каким бы ни был механизм популяционных волн, ясно, что на численность популяции могут влиять одновременно многие факторы. Популяционные волны играют большую роль в ходе микроэволюции. С возрастанием численности популяции увеличивается вероятность появления новых мутаций и их комбинаций. Если в среднем один мутант появляется на 10 тыс. особей, то при возрастании численности популяции в 100 раз общее число мутантов увеличится во столько же раз. После спада волны численности генофонд популяции может уже оказаться иным: часть мутаций может случайно исчезнуть из-за гибели несущих их особей, а частота встречаемости других мутаций может повыситься. Таким образом, популяционные волны сами по себе не вызывают наследственную изменчивость, а только способствуют изменению частот аллелей и генотипов; они являются поставщиком исходного материала для действия естественного отбора.