

Лабораторная работа №11
Тема:Понятие селекции. Организация селекционного процесса с овощными культурами
Цель занятия:Ознакомится с основными понятиями селекции и усвоить организацию селекционного процесса с овощными культурами.
Материалы и оборудование:
1.Схемы,плакаты.
2.Материалы по выполнению лабораторной работы.
Выполнение работы:1. ознакомится и записать в тетрадь основные понятия по селекции овощных культур,систематику растений и понятие осорте;
2.Записать основные селекционные признаки и их классификацию.
3. Изучить способы размножения и биологию цветения,записать основные понятия,
4.Изучить схему и особенности этапов селекционного процесса.
5. Ответить на вопросы.
Тема:Понятие селекции.Организация селекционного процесса с овощными культурами
Селекция — наука и практика выведения новых и совершенствования имеющихся сортов и гибридов сельскохозяйственных растений. Латинское слово «selectio» означает отбор, или выбор, учение о селекции по своей сущности охватывает значительно более широкий круг вопросов, чем отбор. Селекцию можно представить как процесс направленного изменения наследственности растения применительно к условиям внешней
среды и элементам агротехники. По образному выражению академика Н. И. Вавилова, «селекция — это эволюция, направляемая волей человека». Теоретической базой селекции служит генетика, — наука о наследственности и изменчивости организмов. Важное место в селекции растений занимает изучение методов создания исходного материала путем гибридизации, мутагенеза, полиплоидии и других приемов, изменчивости и наследственности признаков и их оценки, методов и направления отбора для получения новых форм, сортов и гибридов.
В селекции большое значение имеет использование факторов внешней среды для выявления таких признаков, как зимостойкость, холодостойкость, засухоустойчивость, иммунитет, и т. д. Вот почему практическая селекция немыслима без участия ряда смежных естественных (физиология растения, биохимия, фитопатология и др.) и отдельных технических наук. Это связано с необходимостью создания сортов, пригодных для машинной уборки, так как роль сорта как одного из главных средств производства возрастает в условиях механизации, химизации и мелиорации сельского хозяйства. Селекционный процесс можно представить как технологию выведения сортов.
Новые сорта овощных культур (капусты, томата, огурца, корнеплодов и др.) должны обладать не только пригодностью к машинной уборке и хорошей транспортабельностью, но и устойчивостью к вирусам, бактериальным и грибным заболеваниям,
вредителям.
Селекционер должен хорошо знать законы наследственности и изменчивости организмов, обоснованно подбирать и включать в селекционный процесс исходный материал, учитывая его потенциальные возможности.
СИСТЕМАТИКА ОВОЩНЫХ РАСТЕНИЙ. ПОНЯТИЕ О СОРТЕ
Систематика. Систематика растений — наука, предметом изучения которой являются многообразие растений, их описание, наименование, классификация и установление системы родственных связей между таксономическими единицами на основе эволюции растительного мира от древнейших примитивных форм до современных.
«Международным кодексом ботанической номенклатуры», принятым XI Международным ботаническим конгрессом в 1969 г., в систематике растений установлены 23 таксона: растительное царство, отдел, подотдел, класс, подкласс, порядок, род, подрод, , секция , подсекция , ряд , подряд , вид , подвид , разновидность ,подразновидность , форма , подформа . Основные таксоны — отдел, класс, порядок, семейство род и вид, остальные служат для обозначения более мелких подразделений. Основная единица ботанической систематики — вид
Ботаники-систематики обычно оперируют крупными таксономическими единицами. Создавая стройную филогенетическую систему растительного мира, они, естественно, ограничиваются определением вида и его места в этой системе. Для селекционера обычно с одним-двумя видами культивируемых или имеющих дело с большим внутривидовым разнообразием наибольший интерес представляет внутривидовая систематика.
Вид- представляет собой сложную популяцию, в которой идет непрерывный мутационный процесс и отбор.
Сорт и сортотип, объединяющий группу близких по многим признакам и характеру хозяйственного использования сортов.
В практической деятельности селекционер оперирует группами растений, которые являются потомствами одного или нескольких более или менее близких растений. В зависимости от их числа, степени генотипического разнообразия, величины гетерозиготности каждого и способа размножения особи каждой конкретной группы могут наследственно отличаться одна от другой в различной степени. В одну группу могут входить очень разнообразные растения, в другую — довольно близкие и в третью — почти идентичные. В зависимости от степени фенотипического и генотипического разнообразия и от способа получения семян такие группы имеют разное селекционное значение и различные названия: популяция, линия, чистая линия, клон и семья. Все это не только внутривидовые, но и часто внутрисортовые единицы.
Популяция — биологически сложившееся сообщество в разной мере родственных особей. Обычно этот термин применяют к сравнительно разнообразным сообществам, например в ботанике по отношению к виду, в растениеводстве — к сортам. В последнем случае часто указывают «сортовая популяция». Типичная популяция — второе гибридное потомство, полученное при скрещивании сильно различающихся растений.
В популяции наблюдается как фенотипическое, так и генотипическое разнообразие особей. Среди сравнительно большого числа составляющих популяцию растений могут одновременно присутствовать очень близкие друг к другу, различающиеся по нескольким или одному признаку, а также растения, которые различаются по многим признакам.
Линия — внешне однородная популяция, выравненность которой поддерживается отбором. Термин «линия» чаще применяют к группе особей селекционного материала, отличающихся от остальных характерным признаком или группой признаков и сохраняющих это отличие в течение ряда поколений. Большинство особей линии гомозиготно по генам,
определяющим характерные для нее признаки, но не исключено наличие гетерозигот по другим признакам, особенно по количественным.
Чистая линия— наиболее выравненный селекционный материал Это потомство, полученное в результате самоопыления гомозиготного растения.
Особи чистой линии однородны как фенотипически,так и генетически.
Естественно, что легче вывести чистую линию при работе с самоопыляющимися растениями, так как они сравнительно мало гетерозиготны и самоопыление для них является нормальным способом размножения. Но чистую линию можно получить и у перекрестноопыляющихся растений, применяя длительный инбридинг. Разработаны методы получения чистых линий удвоением числа хромосом у гаплоидных растений и размножением их самоопылением.
Клон — вегетативное потомство одного растения. Оно обладает большой генотипической однородностью. Практически особи одного клона генетически идентичны между собой и с материнским растением, если только в клетках меристем почек материнского растения, используемых для получения вегетативного потомства, не было соматических мутаций. Наблюдаемые различия между растениями клона имеют модификационный характер и могут быть вызваны как разными условиями выращивания, так и тем, с какой части материнского растения был взят орган вегетативного размножения (черенок, почка, луковица и др.).
Семья — половое потомство, полученное от одного материнского растения. Этот термин применяется независимо от степени гетерозиготности материнского организма. Оно может быть гибридом любого поколения или гетерозиготой перекрестноопыляющегося растения и растением самоопыляющегося сорта, гомозиготность которого не доказана. Семена от материнского растения могут быть получены как в результате самоопыления, так и переопылением с любым числом других растений.
Если обобщить особенности применения перечисленных терминов к тем или иным группам растений в селекционной практике, то потомство от одного растения может быть или семьей, или чистой линией, или клоном. Вегетативное размножение даст клон, самоопыление гомозиготы — чистую линию, а во всех других случаях — семью (рис. 1).
Потомство, полученное от нескольких генотипически различ ных растений, является популяцией. Популяция, обладающая фенотипической однородностью, называется линией.
Понятие о сорте. Сорт (культивар) — выведенная человеком группа хозяйственно ценных по биологическим и морфологи ческим признакам растений, выращиваемых в определенных условиях. Сорт не является определенной систематической единицей, это понятие хозяйственное.
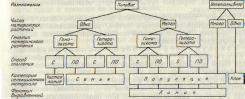
С- самоопыление; ПО- перекрестное опыление
Сорт- популяция с разной степенью наследственной разнокачественностью особей.
Линия – совокупность внешне однородных особей, воспроизводящихся половым путем, размножаемая семенами.
Сорта перекрестноопылющихся растений – пестрые популяции.Они берут начало от нескольких гетерозиготных линий как правило.Бывшая у родительских форм гетерозиготность по большинству генов,определяющих хозяйственно ценные признаки,сохраняется в процессе работы с сортом.только в процессе отбора на выравненность отбираются гетерозиготы по более или менее близким аллелям у перекрестников гомозиготность наблюдается только по отдельным генам,определяющим некоторые морфологически качественные признаки.
Более выравнены сорта самоопыляющихся растений.
Признаки растений
Морфологичесгие признаки:-размер ,форма и окраска луковицы лука.
Наличие волокнистых пучков в корнеплоде свеклы, бобах фасоли, наличие пергаментного слоя в створках бобов гороха, форма и относительный размер ксилемной части корнеплода моркови и др.-анатомические признаки.
В группу физиологических выделяют признаки, характеризующие особенности жизнедеятельности растений (потребность в определенном уровне температуры среды, интенсивности освещения, обеспеченности влагой и питательными веществами, устойчивость к засухе, холоду и др.).
К группе биологических признаков относят особенности роста и развития растений (продолжительность вегетационного периода, реакция на определенную долготу дня, однолетний или многолетний образ жизни и др.).
Качественный и количественный состав сахаров, белков, жиров, витаминов, активность ферментов и другие подобные особенности выделяют в группу биохимических признаков.
Распределение признаков по группам довольно условно. Некоторые признаки можно включить в несколько групп. Например, красная окраска плодов томата — признак морфологический, вместе с тем его можно считать биохимическим, так как он зависит от содержания в тканях определенных пигментов, поэтому строгое закрепление признаков за определенными группами необязательно.
По хозяйственному значению признаки различают: хозяйственно положительные, хозяйственно отрицательные и хозяйственно нейтральные.
К хозяйственно положительным относят признаки, определяющие пригодность растения для культивирования и качество выращиваемого продукта. Так, у капусты к хозяйственно положительным признакам относят высокую плотность кочана, малый размер внутренней кочерыги,белую окраску внутренних листьев, холодостойкость,скороспелость ранних сортов, лежкость кочанов в зимнеевремя и др.
К хозяйственно отрицательным относят признаки, которые затрудняют выращивание растений, делают выращиваемый продукт малоценным (у капусты рыхлость кочана, восприимчивость к киле, плохая лежкость; у томата сильная ребристость плодов; у гороха полегаемость растений и др.). Хозяйственно нейтральные признаки не имеют прямого хозяйственного значения или по крайней мере оно для них пока не установлено (окраска венчика цветка и опушенность листьев у томата, форма стручка у капусты, редиса, репы, окраска семян у салата и др.). Некоторые из нейтральных признаков могут служить показателем однородности сорта, например степень окраски цветка у редиса, форма и размер стручка у капусты и др.
К хозяйственно положительным или отрицательным могут отнесены признаки, входящие в каждую из перечисленных групп (морфологические, анатомические, физиологические, биологические, биохимические и др.). Так, хозяйственная ценность сорта моркови определяется правильной формой корнеплода с гладкой поверхностью и интенсивной окраской (при-таки морфологические), оптимальным соотношением ксилемной и флоэмной части корнеплода (признаки анатомические),высоким содержанием сахаров и каротиноидов (признаки биохимические), устойчивостью к засухе, низким температурам весной (признаки физиологические), дружным прорастанием семян, шинельным периодом покоя при зимнем хранении маточников, плетением и формированием семян при определенной долготе пин (признаки биологические) и другими признаками.
Хозяйственная ценность некоторых признаков не является .абсолютной величиной для всех культур и для разных сортов одной культуры. Она изменяется в зависимости от цели и условий выращивания. Например, крупный размер плодов томата — признак положительный для сортов салатного применения, но отрицательный для сортов, используемых при приготовлении пгльноплодных консервов. Высокая лежкость — признак положительный для поздних сортов капусты, но он не имеет хозяйственной значимости для скороспелых сортов, урожай которых реализуется в середине лета. Жаровыносливость — положительный признак для сортов, выращиваемых в южных районах, но он не имеет хозяйственного значения в северных районах.
Вместе с тем такие признаки, как высокая урожайность, высокое содержание питательных веществ, устойчивость к болезням и вредителям, пригодность к механизированному возделыванию и уборке урожая, являются хозяйственно ценными для большинства культур.
Сорт характеризуется большим числом признаков, из которых каждый может быть обнаружен у какого-то одного или нескольких других сортов данной культуры. Таким образом, один и тот же признак может присутствовать у нескольких разных сортов, и каждый сорт имеет различный набор признаков.
Характерная для сорта группа признаков называется сортовым комплексом признаков. Например, сортовой комплекс признаков томата сорта Маяк 12/20-4 включает следующие признаки: куст обыкновенный, детерминантный, низкорослый, облиственность средняя; листья небольшие, светло-зеленые, гладкие, долек и долечек мало; соцветие простое, плоды плоско-округлые и округлые, гладкие, красные, пяти-девятикамерные; иногда на плодах появляются трещины в виде кольца; скороспелый, урожайность 20—63 т/га; среднеустойчив к засухе, жаре и болезням.
.
В зависимости от характера проявления и наследования различают признаки качественные и количественные.
Качественные признаки показывают наличие или отсутствие определенных особенностей формы, окраски и строения органов, наличие или отсутствие опущенное™, складчатости листа и др. (например, красная и желтая окраска плодов томата, детерминантный и индетерминантный тип строения стебля, черная и белая окраска семян салата, наличие и отсутствие шипов на плодах огурца). Качественные признаки в основном определяют визуально.
Количественные признаки можно легко измерить, подсчитать, определить путем взвешивания и выразить числом (масса кочана, число плодов на растении, длина и ширина листа, продолжительность вегетационного периода). Принципиальных различий между качественными и количественными признаками нет. Такое деление признаков принято в селекционной практике в связи с тем, что они по-разному учитываются и несколько иначе генетически определяются.
Качественные признаки определяются сравнительно небольшим числом генов, которые действуют в комплексе. В результате создается впечатление, что качественный признак определяется действием одного, иногда двух-трех так называемых главных генов. В гибридных, а также в инбредных потомствах от гетерозигот расщепление по качественным признакам подчиняется закономерностям, установленным Менделем. Проявление количественного признака определяется большим количеством генов, число которых достигает одной-двух сотен. При этом каждый из них незначительно влияет на проявление признака, в связи с чем долю действия отдельного гена выявить практически невозможно. Взаимоотношения между разными генами, определяющими один и тот же признак, и аллелями одного гена могут быть самыми разнообразными.
В гибридных потомствах по каждому гену, определяющему количественный признак, наблюдается менделевское расщепление, но так как их очень много и действие каждого незначительно, то в целом расщепление во втором гибридном потомстве по количественным признакам приближается к нормальному распределению.
Качественные признаки почти не изменяются под влиянием условий среды, например окраска зрелых плодов томата остается красной в различных условиях, типичное строение шипов на плодах огурца сохраняется в любых условиях выращивания. Количественные же признаки очень чутко реагируют на изменения условий выращивания, например масса кочана у капусты и салата, корнеплода у моркови, свеклы и редьки, плодов у томата будет различной при выращивании одного и того же сорта в разных условиях.
Перечисленные различия между качественными и количественными признаками при всем их обилии нисколько не умаляют высказанного положения, что между ними нет принципиальных различий. Подтверждением этому может служить такой пример. Но втором гибридном потомстве, полученном от скрещивания сорта редиса, имеющего красноокрашенный корнеплод, с сортом с белым корнеплодом, расщепление по признаку наличия фиолетовой, красной и белой окраски определяется двумя комплементарно действующими главными генами и равно 9:3:4. Однако степень окрашивания, которая изменяется у красноокрашенных форм от темно-красной до бледно-розовой и у фиолетовоокрашенных от темно-фиолетовой до сиреневой, наследуется как количественный признак. Следовательно, проявление одного и того же признака имеет и качественный, и количественный характер.

Рис. 2. Изменчивость корнеплода моркови сорта Шантенэ в зависимости от площади питания (см)
Изменения факторов среды,или площади питания Например, у репчатого лука сильнее реагируют такие признаки, как диаметр и высота луковицы, слабее — толщина чешуи (табл. 1).
1. Влияние площади питания на морфологические признаки репчатого лука
Показатель |
Площадь питания, см |
Г Коэффициент варьирования |
|
|
|
|
|
|
4X4 |
8X8 |
12X12 |
16X16 |
20X20 |
|
|
Сорт Мячковский |
||||||
Диаметр, мм |
33 |
50 |
57 |
72 |
76 |
30 2 |
Высота, мм |
29 |
36 |
41 |
45 |
44 |
33 8 |
Зачатковость |
1,01 |
1,32 |
2,01 |
1,97 |
1,99 |
28 0 |
Толщина |
3,6 |
4,6 |
5Д |
5,3 |
5,6 |
16,1 |
чешуи, мм |
|
|
|
|
|
|
Сорт Ростовский репчатый |
||||||
Диаметр, мм |
22 |
33 |
46 |
55 |
57 |
35 0 |
Высота, мм |
29 |
39 |
43 |
46 |
47 |
40,7 |
Изменчивость признаков в онтогенезе. В селекционной практике с изменчивостью признаков, определяемой онтогенетическим состоянием растения, встречаются постоянно, когда необходимо охарактеризовать растения по морфологическим признакам их органов. Для точной оценки необходимо знать, как изменяются признаки в процессе формирования отдельного органа и в онтогенезе всего растения.
Как известно, высшие растения представляют собой довольно сложные системы взаимосвязанных повторяющихся (метамерных) органов: корневая система, листья, узлы и междоузлия стеблей, цветки и плоды. Овощные растения характеризуются еще и тем, что многие из них имеют одиночные (неметамерные) сильно гипертрофированные органы: корнеплод, луковицу, кочан, стеблеплод и головку, ради которых их выращивают.
Каждый орган в процессе роста и развития претерпевает изменения. В соответствии с этим изменяются его признаки. Вместе с тем метамерные органы, закладываясь последовательно в точках роста побегов, проходят этапы формирования в разное время жизни растения, на разных этапах онтогенеза. Вследствие этого внутренние условия, определяющие их рост и развитие, а также степень выражения их признаков будут различными. Следовательно, при анализе признаков растения нужно учитывать их изменчивость как в онтогенезе органа, так и всего растения.
Наблюдения показали, что в процессе развития органа признаки изменяются различно. Особенно подвержены колебаниям признаки, характеризующие линейные размеры органа (длина и ширина листа, побега, плода, корнеплода, стеблеплода и др.). Значительно изменяется форма органа. Например, форма листа розетки салата вначале бывает более округлой, а по мере роста листа она становится округло-уплощенной. При этом ширина листа может быть больше длины более чем в 1,5 раза. Значительно изменяется форма корнеплода. Так, у редиса, редьки, свеклы и других культур в процессе роста форма корнеплода изменяется от удлиненной через овальную к округлой и даже округло-плоской у отдельных сортов У некоторых сортов моркови молодые корнеплоды имеют овально-удлиненную форму, затем коническую, в зрелом состоянии цилиндрическую.
СПОСОБЫ РАЗМНОЖЕНИЯ И БИОЛОГИЯ ЦВЕТЕНИЯ
Начальным этапом селекционной работы с любой овощной культурой является изучение способа ее размножения и особенностей биологии цветения, если она размножается половым способом. Свойственные каждой культуре особенности размножения в значительной мере определяют возможность выбора того или иного селекционного метода.
При работе с самоопыляющимися растениями селекционер применяет иные способы воздействия на состав селекционных популяций и защиты растений от подмешивания постороннего генетического материала, чем при селекции перекрестноопыляющихся. Зная биологию цветения растений, можно выполнять работы по кастрации и опылению в оптимальные фазы развития цветка, при благоприятном сочетании факторов внешней среды. Способ опыления растений определяет возможность применения того или иного типа изоляторов и выбор расстояний между участками размножаемых растений. От способа размножения и характера опыления цветков зависят также скорость закрепления нужных признаков в селектируемых популяциях и быстрота размножения нового сорта при его семеноводстве. Большинство овощных растений размножается половым способом, и только у некоторых из них он сочетается с бесполым (лук репчатый, спаржа, эстрагон и др.).
Бесполое размножение. Апомиксис («апо» — отсутствие, «миксис» — смешивание, слияние половых клеток или наследственности), или бесполое размножение, — это такой способ размножения, при котором не происходит оплодотворения. Новые особи развиваются из вегетативных почек, образовавшихся в пазушных луковичках-зубках, и дочерних околокорневых луковичек у чеснока, в воздушных луковичках-бульбочках у многоярусного лука и чеснока, на искусственно отделяемых частях корневищ у спаржи, эстрагона, щавеля и др., из почек, образовавшихся на корневых черенках у хрена, и т. д. Бесполое размножение у овощных растений распространено сравнительно мало, чаще его используют у декоративных, ягодных и плодовых растений.
Бесполое размножение может происходить и в генеративных органах. Форму апомиксиса, при которой зародыш развивается из неоплодотворенной яйцеклетки, называют партеногенез) Иногда вследствие дегенерации яйцеклетки зародыш развиваается из других клеток зародышевого мешка — синергид и антипод. Такие формы апомиксиса называют синергидной и антиподной апогамией. При партеногенезе и апогамии формирующийся зародыш семени имеет гаплоидный набор хромосом, так как он развивается в результате деления клеток, имеющих гаплоидные ядра. Часто эти формы апомиксиса сочетаются с нарушением хода мейоза, в результате которого не происходит расхождения хромосом, и клетки зародышевого метка имеют ядро с двойным числом хромосом. Вследствие этого образующиеся в результате партеногенеза и апогамии зародыши бывают диплоидными. У некоторых растений наблюдается формирование зародыша из клетки нуцеллуса, такую форму апомиксиса называют апоспорией. При апоспории зародыш имеет диплоидный набор хромосом.
При всех перечисленных формах апомиксиса, происходящего в генеративных органах, из инициальной клетки формируется зародыш, а из тканей семяпочки — покровы семени. В конечном итоге растение размножается семенами. Однако из этих семян вырастают растения с идентичной материнскому растению наследственностью.
Среди овощных растений не обнаружено форм, обладающих той| или иной формой апомиксиса в генеративных органах, хотя не исключено его присутствие как отклонение от нормы в отдельных семяпочках. Наиболее часто партеногенез наблюдается при опылении рыльца пыльцой с растений другого вида или рода при обработке его физиологически активными веществами. В этом случае в результате возбуждения яйцеклетки формируется партеногенетический зародыш, который чаще бывает гаплоидным.
Такое явление, когда пыльца выступает только в роли стимулятора деления яйцеклетки вследствие опыления или прорастании пыльцевой трубки в ткань пестика, а ее наследственные элементы не включаются в ядро яйцеклетки, называется псевдогамией. Например, при опылении пестика капусты пыльцой индау (Егuса sativа) пыльцевые трубки проникают в ткань пестика, достигают микропиле, но оплодотворение не производят. Из возбужденной яйцеклетки формируется гаплоидный заромыщ семени капусты.
Вегетативное размножение вносит определенную специфичное и, в селекционную работу с растениями. Отрицательная сто- рона вегетативного размножения состоит в том, что коэффициент размножения в этом случае очень мал по сравнению с семенным. Вместе с тем при работе с вегетативно размножаемыми растениями отпадает необходимость проведения в течение ряда поколений отбора с целью выравнивания селекционных популяций.
Половое размножение. Большинство овощных культур размножают половым способом, что позволяет от отдельного растения получить большое количество особей потомства. Однако коэффициент размножения имеет разную величину у представителей разных видов растений. Так, если у гороха можно получить с одного растения несколько десятков семян, в лучшем случае немногим более сотни, то у кочанной капусты и других растений — несколько десятков тысяч. Как и другие признаки растений, коэффициент размножения зависит от условий выращивания, его величина может варьировать в широких пределах.
Половое размножение в то же время значительно затрудняет выравнивание селекционных популяций, так как в потомстве от гетерозиготных растений, с которыми селекционеру, как правило, приходится иметь дело, наблюдается очень большое разнообразие особей вследствие генетического расщепления.
По характеру опыления размножающиеся половым путем растения разделяют на две группы: самоопыляющиеся и перекрестноопыляющиеся.
К самоопыляющимся относят такие растения, у которых наблюдается опыление рыльца пыльцой и оплодотворение яйцеклеток спермиями из пыльцевых трубок того же цветка. Самоопыление, или автогамия, таким образом обеспечивает получение потомства, разнообразие которого определяется только степенью гетерозиготное™ материнского организма. У разных культур самоопыление зависит от особенностей строения цветка и функционирования его органов.
Так, у гороха пыльники и пестик находятся в лодочке цветка в непосредственной близости одни к другому, созревают почти одновременно. Еще в фазе бутона пыльники вскрываются и пыльца полностью обволакивает столбик и рыльце. Это обеспечивает высокую гарантию опыления своей пыльцой. Опыление чужой пыльцой возможно только в редких случаях, когда лодочка венчика повреждается грызущими насекомыми до вскрытия пыльников или при посещении шмелями и другими насекомыми цветков с маложизненной пыльцой.
В цветках салата пыльники вскрываются к моменту выхода рыльца из колонки пыльников наружу. Рыльце продвигается около лопнувших пыльников, пыльца попадает на края лопастей рыльца, которые к моменту выхода наружу и развертывания бывают уже опылены. Опыление чужой пыльцой хотя и происходит с помощью посещающих соцветия насекомых, но оно во
времени отстает от самоопыления. В результате образование семян от опыления пыльцой с других растений наблюдается очень редко.
У томата пыльники, соединяясь между собой выростами эпипермальных клеток, образуют колонку, внутри которой находится столбик пестика. Рыльце расположено на одном уровне с концами пыльников в непосредственной близости от них. При покрытии пыльников пыльца легко попадает на рыльце и оплодотворяет завязь. Самоопыление гарантируется не только строением цветка, но и тем, что цветки томата сравнительно редко посещают насекомые. Частичное образование семян от опыления другими растениями наблюдается иногда в жаркие годы, при посещении цветков шмелями, трипсами и другими насекомыми.
В несколько большей мере опыление пыльцой других растений наблюдается у других представителей семейства Пасленовые — перца и баклажана, у которых цветки хотя и имеют почти одинаковое строение, но рыльце бывает расположено несколько выше тычинок, образующих довольно рыхлый конус.
Кроме особенностей строения цветка, на вероятность опыления чужой пыльцой существенно влияют условия внешней среды что в мере определяется как строением, так и размерами цветков.
Преимущество самоопыления с точки зрения селекции состоит в том, что при выравнивании селекционных популяций нет необходимости тщательно изолировать растения, что облегчает организацию селекционной работы. Можно также выращивать различные группы растений на одном участке сравнительно близко одну от другой.
Самоопыление определяет и соответствующий характер поведения гибридных и других созданных селекционером гетерозиготных популяций. При размножении в течение ряда поколений происходит довольно быстрое накопление гомозигот в результате естественного расщепления гетерозигот в естественных условиях новые комбинации генов могут возникнуть только вследствие мутационного процесса. Появившаяся мутация сохраняется только в пределах потомства того растения, у которого она возникла.
Популяции самоопыляющихся сортов представляют собой сообщества в разной мере родственных гомозиготных по одним и гетерозиготных по другим признакам линий. Отбор, проводимый человеком в пределах сорта самоопыляющейся культуры, приводит только к количественному изменению соотношения различных генотипов, а не к возникновению новых комбинаций генов.
К перекрестноопыляющимся растениям относятся большинство овощных культур, у них оплодотворение яйцеклетки возможно спермием пыльцы другого растения данного вида. У перекрестноопыляющихся, или ксеногамных, растений имеются разнообразные приспособления, которые в той или иной мере препятствуют попаданию своей пыльцы на рыльце от автогамного (с того же цветка) или гейтеногамного (с других цветков того же растения) опыления. У шпината, спаржи и других растений мужские и женские цветки находятся на разных растениях. Перекрестное опыление гарантируется наличием у растения только одного пола — женского или мужского. Особи разного пола находятся в популяции примерно в равном количестве. У сахарной кукурузы, огурца, кабачка, патиссона и других тыквенных растений женские и мужские органы находятся в разных цветках, что уменьшает возможность попадания своей пыльцы на рыльце, но не исключает этого полностью. У сахарной кукурузы, кроме того, мужские и женские соцветия пространственно разобщены и возможность перекрестного опыления усиливается благодаря дихогамии — разновременному созреванию тычинок и пестиков. Цветение мужского соцветия на растении начинается на 1—2 дня раньше, чем появляются рыльца из обертки початка.
Дихогамия препятствует самооплодотворению и у растений с обоеполыми цветками — моркови, свеклы, лука, капусты, репы и др. У моркови и лука пыльники созревают, вскрываются и освобождаются от пыльцы на 2—4 дня раньше того момента, когда рыльце становится восприимчивым к пыльце. Такое явление раннего созревания пыльников носит название протерандрии. У капусты, репы, редиса и других растений семейства Крестоцветные* (Капустные) рыльце созревает еще в фазе бутона, а пыльники вскрываются через несколько часов после раскрытия цвет-
СХЕМА СЕЛЕКЦИОННОГО ПРОЦЕССА
Движущими факторами селекции, как и эволюции, являются изменчивость, наследственность и отбор. Однако проявление этих факторов, их активность и направленность в селекции несхожи с наблюдаемыми в дикой природе. Селекция отличается от естественного эволюционного процесса тем, что при необходимости, применяя мутагенные факторы, селекционер может в сотни и тысячи раз увеличить мутационную изменчивость организмов. Зная природу наследственности и учитывая особенности способа размножения, селекционер имеет возможность объединить в результате гибридизации комплексы наследственных факторов разных организмов и тем изменить ход и направление изменчивости. И, наконец, очень важное отличие селекции состоит в том, что наряду с естественным в селекционной работе особую роль приобретает искусственный отбор, проводимый человеком. Часто направления естественного и искусственного отборов не совпадают. Особенно ярко это выражено в селекции овощных растений, большинство из которых имеют сильно гипертрофированные органы (кочан, партенокарпические плоды и т. д.), строение которых препятствует нормальной их жизнедеятельности или требует значительных затрат продуктов фотосинтеза в ущерб семенной продуктивности. Морфобиологическая структура большинства овощных растений является следствием двух противоположно направленных факторов — естественного и искусственного отборов и находится в состоянии неустойчивого равновесия.
Сущность селекции состоит в улучшении старых и создании новых форм растений. Во время селекционной работы селекционеру приходится выполнять различные приемы, проводить исследования, применять разные способы создания популяций. Весь комплекс мероприятий, проводимых селекционером от начала работы до создания такого селекционного материала, который в качестве нового сорта может быть включен в станционное и затем государственное сортоиспытание, называют селекционным процессом.
В зависимости от цели селекционной работы, длительности цикла развития, способа размножения и других биологических особенностей растений мероприятия, выполняемые селекционером в процессе его работы, могут быть весьма разнообразными. Комплекс выполняемых в процессе селекции мероприятий в соответствии с тем, с какой целью их проводят, какие методы при этом применяют, можно разделить на три этапа: подбор и создание исходного материала для отбора, отбор и испытание.
I этап. На первом этапе в соответствии с поставленной задачей селекционер, мысленно представляя будущий сорт, анализирует многообразие выращиваемых в данной зоне местных и селекционных, а также инорайонных и зарубежных сортов для обнаружения форм, наиболее ему соответствующих. Если в каких-то сортах встречаются формы, близкие к намеченному образцу, то они служат исходным материалом для селекционной
работы (рис. 3).
При отсутствии в изучаемом материале форм, близких к намеченному образцу, или обнаружении отдельных требуемых признаков у представителей разных сортов селекционер искусственно создает популяции исходного материала селекции.
Наиболее эффективный и широко применяемый способ создания популяций исходного материала — гибридизация. Обнаружив требуемые для нового сорта признаки в разных сортах и скрестив последние, селекционер в гибридных организмах объединяет наследственность двух сортов. В полученном от такого растения потомстве он имеет возможность наблюдать большое многообразие сочетаний различных признаков, обусловленное различными комбинациями наследственных факторов этих сортов. У размножаемых семенами самоопыляющихся растений в качестве исходного материала используют обычно второе и следующие гибридные потомства. При скрещивании гетерозиготных перекрестноопыляющихся растений большое разнообразие форм наблюдается уже в первом гибридном поколении, поэтому его, а также второе поколение используют в качестве исходного материала селекции. Если обнаруженные в гибридных популяциях формы недостаточно близки к намеченной селекционером цели,
1-этап Поиск и создание исходного материала селекции
Дикорастущие растения |
|
|
|
Сортобые популяции |
Селекционные |
Местные |
|
Инорайонные |
|
|
|
Гибридные популяции |
Внутривидодые |
Межвидобые |
|
|
|
Полученные популяции |
Мутантов |
Полиплоидов |
|

ПерВое поколение (2-этап) |
Оценка растений, исходной популяции и отделение лучших для размножения |
|
Размножение, или создание популяций следующего поколения |
||
1 |
|
|
Второе поколение (2-этап) |
Оценка растений созданных популяций и Выделение лучших для размножения |
|
Размножение, или создание популяций следующего поколения |
||
и т. д. (до создания популяции., отвечающей
целям селекционной работы)
Ш этап. Испытание
Контрольное
Конкурсное, или станционное
Государственное Рис .3 Схема селекционного процесса
Гибридизация не всегда обеспечивает возможность получения требуемого исходного материала. Во-первых, потому что в пределах сортового многообразия может отсутствовать признак, необходимый для нового сорта, во-вторых, некоторые ценные признаки могут быть сильно связаны с отрицательными признаками, а кроссинговер между ними мало вероятен.
Для получения значительных изменений, например таких, которые у данной культуры не наблюдаются, или, наоборот, для изменения одного признака при сохранении существующего ком-пнгкса в целом применяют создание популяций исходного материала с помощью мутагенных факторов. Для этого воздействуют физическими или химическими мутагенами на растения наиболее перспективных сортов. Полученное многообразие форм может служить исходным материалом для селекции. Но наиболее часто возникших в результате воздействия мутагенными факторами популяций выделяют наиболее интересные растения, скрещивают их с лучшими сортами, а гибридные потомства используют в качестве исходного материала. Иногда для создания популяции исходного материала воздействуют на растения колхицином и дру-1пми веществами, вызывающими кратное увеличение наборов хромосом в клетках, или явление полиплоидии. Возникшие в результате такого воздействия популяции также отличаются определенной специфичностью проявления признаков.
Из перечисленных способов создания популяций исходного материала каждый может быть использован отдельно или в сочетании с другими. Выбор того или иного способа определяется специфичностью поставленной цели, биологическими особенностями растений, эрудицией и интуицией селекционера и теми возможностями, которыми он располагает.
II этап. Цель этого этапа (отбора) — выделение из популяции исходного материала наиболее близких к намеченному образцу растений и создание на базе их наследственности новых популяций, все особи которых будут обладать требуемыми признаками. Па завершение отбора обычно требуется 5—7 поколений. Отбор включает два мероприятия, которые проводят поочередно в каждом поколении: оценку растений и создание популяций следующего поколения.
В каждом поколении отбора оценку растений и выделение наиболее ценных в качестве родоначальников будущих потомств можно выполнить различными методами, специфичность которых определяется числом и значимостью оцениваемых признаков, а также способом оценки каждого признака. Например, ценность анализируемых растений можно определять как по отдельному признаку, так и по комплексу признаков. Один и тот же признак можно оценить несколькими способами. Например,продуктивность растения можно определить как визуально, так и взвешиванием. В первых поколениях отбора, когда материал слишком разнообразен и многочислен, обычно применяют легко выполнимые способы оценки. В более поздних поколениях, когда селекционный материал достигает определенной выравненность и насыщенности ценными растениями, а разница между образцами нивелируется, применяют более сложные и точные анализы.
Различают несколько методов отбора. Их названия и способы проведения определяются особенностями создания популяций следующего поколения. Разнообразие методов отбора, а 'точнее методов получения популяций следующего поколения, определяется тем, сколько потомств вошло в состав каждой популяции (от многих, от небольшого числа растений или от одиночных растений), насколько ограниченна была возможность переопыления у перекрестноопыляющихся растений, а также тем, когда проводится оценка растений (до цветения или после него).
В ходе селекционной работы способы оценки растений и методы создания популяций следующих поколений в случае необходимости могут быть изменены в зависимости от изменения качества и количества селекционного материала. Их правильное сочетание в значительной мере определяет успех селекционной работы.
III этап. Этот этап селекционного процесса включает проведение различных по объему и сложности испытаний полученного в результате селекционной работы материала. Различают предварительные и конкурсные (станционные) испытания.
Контрольные вопросы и задания
Дайте определение селекции.
2. Назовите имена выдающихся ученых, работавших в области генетики, селекции и семеноводства овощных и других культур.
3. Расскажите о вкладе отечественных ученых в развитие генетики, селекции и семеноводства овощных и других культур.
4. Назовите типы бесполого размножения растений и охарактеризуйте их значение в селекции.
5. В чем заключаются преимущества и недостатки селекционной работы с перекрестноопыляющимися и самоопыляющимися растениями?
6. Расскажите об особенностях строения и функций цветков перекрестноопыляющихся растений, препятствующих самоопылению.
7. Охарактеризуйте состав сортовых популяций самоопыляющихся и перекрестноопыляющихся растений
8. На какие этапы подразделяют селекционный процесс?
9. Какие мероприятия выполняют на каждом этапе?
10. Почему направления естественного и искусственного отборов в селекции овощных растений не совпадают?
11. Какой способ создания исходного материала наиболее широко применяют в селекции овощных культур?
Лабораторная работа №12
Тема: Особенности селекции капусты и изучение варьирования признаков
Цель занятия: Ознакомится с основными направлениями селекции капусты и изучить варьирование признаков растений.
Материалы и оборудование:
1.Схемы,плакаты.
2.Материалы по выполнению лабораторной работы.
Выполнение работы:
1. Ознакомится и записать в тетрадь основные положения по селекции культуры капусты.
2.Записать основные селекционные и морфологические признаки .
3.Изучить схему и особенности этапов селекционного процесса.
4. Ответить на вопросы
КАПУСТНЫЕ РАСТЕНИЯ
Капуста — одна из основных овощных культур. В России ее выращивают повсеместно, занимает она около 30 % общей площади овощных культурна в Нечерноземной зоне и Сибири — до 40-50 %. Широкому распространению капусты способствовали ее холодостойкость, высокая урожайность и значительная питательная ценность в сочетании с хорошими вкусовыми качествами.