

Рецепция андрогенов
Импульсные выбросы из гипоталамуса гонадотропин-рилизинг гормона стимулируют выделение гипофизом лютеинизирующего и фолликулостимулирующего гормонов, которые называют гонадотропинами. Лютеинизирующий гормон стимулирует производство тестостерона клетками Лейдига и способствует развитию яичек. Фолликулостимулирующий гормон совместно с тестостероном регулирует сперматогенез и созревание сперматозоидов. Кроме того, фолликулостимулирующий гормон усиливает реактивность клеток Лейдига к лютеинизирующему гормону, индуцируя появление ЛГ-рецепторов на клеточных мембранах. Поскольку секреция гонадотропинов носит импульсный характер с наибольшим пиком секреции в утренние часы, то и секреция тестостерона также имеет циркадный ритм, с повышением секреции в 6.00-8.00 часов и понижением в вечерние часы (20.00-22.00). Тестостерон регулирует выделение лютеинизирующего и фолликулостимулирующего гормонов посредством отрицательной обратной связи гипоталамуса и гипофиза. После секреции яичками тестостерон в кровеносном русле взаимодействует по принципу обратной связи с гипоталамо-гипофизарной системой и, таким образом, тормозит выделение лютеинизирующего гормон. Это классическое физиологическое взаимодействие служит для поддержания сывороточного тестостерона на необходимом уровне. Если уровень сывороточного тестостерона падает, ниже определенного, происходит стимуляция гипофиза, который секретирует лютеинизирующий гормон, тот, в свою очередь, стимулирует выделение яичками тестостерона, пока содержание тестостерона в сыворотке крови не достигнет необходимого значения. Наоборот, если содержание сывороточного тестостерона превышает необходимый уровень, пониженное выделение лютеинизирующего гормона снижает секрецию яичками тестостерона до тех пор, пока концентрация тестостерона в сыворотке крови не снизится до нужного значения.
Регуляция спермообразующей функции является более сложной и значительно менее понятной, чем регуляция уровня тестостерона. Фолликулостимулирующий гормон индуцирует запуск и процесс сперматогенеза, действуя через клетки Сертоли. Одновременно фолликулостимулирующий гормон стимулирует высвобождение гормона ингибина из клеток Сертоли в общий кровоток. Циркулирующий ингибин тормозит выделение фолликулостимулирующего гормона по принципу обратной связи. Если интенсивность сперматогенеза уменьшается, то продукция ингибина также уменьшается и, таким образом, эффект обратной связи на гипофиз будет меньше. Это снижение секреции ингибина приводит к увеличению выделения фолликулостимулирующего гормона, который опять усиливает стимуляцию сперматогенеза через клетки Сертоли. Однако не все аспекты этой обратной связи (фолликулостимулирующий гормон - ингибин - сперматогенез) убедительно доказаны. Гипофиз выделяет также пролактин. Для поддержания максимальной стероидогенной активности клеток Лейдига в присутствии лютеинизирующего гормона пролактин необходим. Он увеличивает число рецепторов к лютеинизирующему гормону. Однако повышенный уровень пролактина оказывает отрицательное влияние на секреторную функцию клеток Лейдига.
Ранее считали, что гормоны влияют только на клетки-мишени, удаленные от эндокринных желез - эндокринный механизм действия гормонов. В настоящее время известно, что гормоны могут действовать на соседние клетки эндокринной железы - паракринный механизм. А также даже на клетки, где они сами вырабатываются - аутокринный механизм. Резистентными к гормону могут быть любая из клеток-мишеней или все эти клетки.
Каждый стероидный гормон действует через свой специфический рецептор, расположенный в ядре клеток-мишеней. Гормон проникает в клетку пассивно и находит свой рецептор в комплексе с так называемыми белками теплового шока, которые защищают клетку от различных стрессовых воздействий (высокой температуры, гипоксии, изменения рН и т.д.). После того как гормон соединяется со своим рецептором, происходит изменение пространственной трехмерной структуры последнего, в результате чего рецептор освобождается от белка теплового шока.
Затем запускается сложный каскад реакций, включающий фосфорилирование, димеризацию (удвоение комплекса гормон-рецептор), перемещение его в ядро, взаимодействие со специфической последовательностью ДНК, активацию РНК-полимеразы и усиление транскрипции регулируемого гена. В результате изменяется синтез белков, ответственных за биологические эффекты гормонов.
Взаимодействие стероидного гормона и его рецептора находится под контролем особых ядерных белков: корепрессоров и коактиваторов. Эти белки играют, вероятно, основную роль в тканеспецифичности селективных модуляторов. Коактиваторы способствуют взаимодействию рецепторов с ДНК, а корепрессоры удерживают рецепторы в неактивном состоянии. Если лиганд является агонистом рецепторов, происходит взаимодействие комплекса лиганд-рецептор со специфическим коактиватором. Если лиганд является антагонистом рецепторов, взаимодействия с коактиватором не происходит.
В целом на активность стероидных рецепторов в различных тканях влияют несколько факторов:
• концентрация стероидных рецепторов;
• концентрация лиганда;
• функция лиганда (агонист, частичный агонист, антагонист)
• концентрация и тип коактиваторов и корепрессоров;
• фосфорилирование ядерных рецепторов
Рецепторы лютеинизирующего гормона и хорионического гонадотропина - одни из самых крупных рецепторов. Каждый рецептор лютеинизирующего гормона и хорионического гонадотропина состоит из внеклеточного домена, трансмембранного домена, содержащего 7 субъединиц, и внутриклеточного домена. Структура рецептора фолликулостимулирующего гормона изучена хуже. Обнаружено и клонировано несколько генов, кодирующих рецепторы лютеинизирующего гормона и хорионического гонадотропина. Все эти гены локализуются на 2-й хромосоме. Ген рецептора фолликулостимулирующего гормона также клонирован; он локализуется на 14-й хромосоме. Рецепторы лютеинизирующего гормона и хорионического гонадотропина и рецепторы фолликулостимулирующего гормона обнаружены не только в половых железах, но и во многих других органах.
Рецепторы андрогенов также принадлежат к семейству внутриклеточных рецепторов. Эти рецепторы локализуются в цитоплазме и включают гормонсвязывающий домен, ДНК-связывающий домен и домен, активирующий транскрипцию гена-мишени. Рецептор андрогенов- это гликопротеид, содержащий 910 аминокислот. Ген рецептора андрогенов расположен на X-хромосоме и состоит из 100000 нуклеотидов. Рецептор андрогенов способен связывать как тестостерон, так и другие андрогены. Аффинность связывания максимальна для дигидротестостерона, несколько ниже - для тестостерона и минимальна - для андростендиона. Андрогены и функционирующий андрогеновый рецептор (АР) являются ключевыми в развитии и поддержании мужского фенотипа и сперматогенеза. Мутации гена андрогенового рецептора (АР) вызывают разнообразные дефекты, связанные с потерей чувствительности к андрогенам. Это проявляется в виде различных нарушений - от полной феминизации до фенотипических мужчин, страдающих бесплодием. Резистентность к андрогенам - распространенная причина мужского псевдогермафродитизма у лиц с кариотипом 46 ХУ.
Андрогены осуществляют свои эффекты преимущественно через классический ядерный рецептор, влияющий на транскрипцию генов. Кроме того, андрогены, как и другие классы стероидных гормонов, оказывают быстрые негеномные эффекты в результате взаимодействия с классическими рецепторами стероидов или рецепторами, сопряжёнными с G-белками, локализованными в плазматической мембране. Современные представления о действии андрогенов в клетках-мишенях приведены на рисунке.
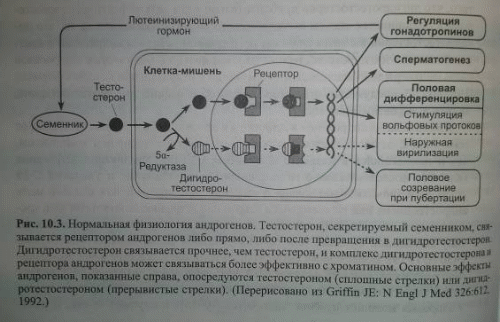
Среди основных функций андрогенов - регуляция секреции гонадотропинов гипоталамо-гипофизарной системой, стимуляция сперматогенеза, формирование мужского фенотипа во время половой дифференцировки и стимуляция полового созревания при пубертации. В клетке тестостерон может превращаться в дигидротестостерон под действием стероид-5-редуктазы 1 или 2; стероид-5-редуктаза 2 является преобладающим изозимом в большинстве тканей-мишеней для андрогенов. Тестостерон или дигидротестостерон затем связывается с белком рецептора андрогенов. Рецептор андрогенов кодируется геном на длинном плече X-хромосомы и имеет размер приблизительно 110000 Да. Он сходен с другими членами семейства рецепторов стероидов/тиреоидных гормонов/ретиноидов с индивидуальными гормонсвязывающим, ДНК-связывающим и функциональным доменами.
В несвязанном состоянии рецептор андрогенов находится преимущественно в ядре. Связывание андрогена с гормонсвязывающим доменом рецептора с образованием гормон-рецепторных комплексов является необходимым условием для реакции трансформации, при которой гормон-рецепторные комплексы приобретают повышенное сродство к ядерным компонентам. После этого ДНК-связывающие домены рецепторов связываются с андрогенчувствительными элементами (АЧЭ) промоторной области генов-мишеней, активируя транскрипцию.
На основании исследований на человеке и животных представляется, что тестостерон-рецепторный комплекс ответственен за регуляцию гонадотропинов, стимуляцию сперматогенеза и вирилизацию вольфовых протоков во время половой дифференцировки, тогда как дигидротестостерон-рецепторный комплекс ответственен за наружную вирилизацию во время эмбриогенеза и за большинство мужских вторичных половых признаков у взрослых. Концентрация рецептора андрогенов является наибольшей в добавочных органах мужской репродуктивной системы, рост которой зависит от андрогенов. В других тканях, таких как скелетная мышца, печень и сердце, содержание рецептора ниже. В семеннике рецепторы андрогенов находятся и в изолированных клетках Сертоли, и в интерстициальных клетках. Неясно, можно ли на основании присутствия рецептора андрогенов идентифицировать ткань как андрогенчувствительную, хотя рецепторы андрогенов находятся в большем количестве тканей, чем те, о которых известно, что они отвечают на андрогены. Мутации одного гена у человека и животных показывают, что и тестостерон, и дигидротестостерон связываются одним рецептором андрогенов и что рецепторный белок кодируется локусом на X-хромосоме. Для нормального андрогенного действия, по-видимому, необходимо образование дигидротестостерона, несмотря на то, что имеется всего лишь один рецептор. В случае рецептора андрогенов человека частичным объяснением этой необходимости в дигидротестостероне может служить в несколько раз более высокое сродство рецептора к дигидротестостерону по сравнению с тестостероном. Дигитротестостерон может также более эффективно облегчать трансформацию рецептора для трансактивации. Таким образом, различие во взаимодействии двух гормонов с рецептором андрогенов может служить в качестве механизма усиления действия андрогенов в некоторых тканях-мишенях.