

9.4. Терморецепторы
У млекопитающих и многих других видов животных есть специфичные тепловые и холодовые терморецепторы, не чувствительные к нетермическим стимулам [2, 6, 7, 9, 11, 12, 13, 25, 30]. Холодовые обычно иннервируются волокнами группы III (Αδ), а тепловые - группы IV (С). Гистологически на окончаниях холодовых рецепторов выявляется специальная структура; для тепловых гистологической специализации пока не обнаружено, - должно быть, они относятся к категории свободных нервных окончаний.
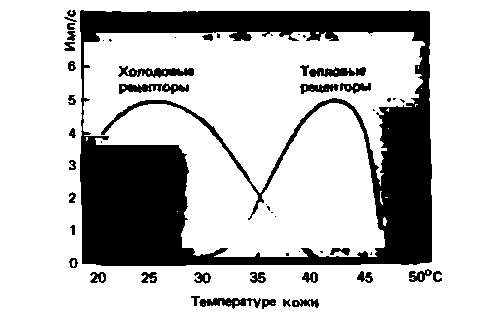
Реакции при постоянной температуре кожи. На
рис. 9.12 показана частота импулъсации терморецепторов в зависимости от постоянной температуры кожи. Это статическая частота импульсации у тепловых рецепторов возрастает при переходе примерно от 30 до 43 оС, а затем резко падает; в пороговой области ощущения боли от горячего (приблизительно 45 °С) тепловые рецепторы обычно невозбудимы. Частота импульсации холодовых рецепторов повышается по мере падения температуры от 33-40 °С и достигает максимума при 17-26 оС (у разных рецепторов). Из-за этого максимума каждая частота разрядов связана с двумя температурами, но тем не менее ЦНС может правильно распознавать холод и на отрезке ниже 17 оС. Как - еще неизвестно [7]. Возможно, для этого нужна динамическая реакция Холодовых ре-
|
Рис. 9.12. Статические характеристики Холодовых и тепловых рецепторов в коже обезьяны. Кривые показывают среднюю частоту импульсации в нескольких афферентах каждой популяции рецепторов в зависимости от неизменной температуры тела. Импульсация холодовых рецепторов снова возрастает при температурах выше 45 °С. Данные получены для одиночных нервных волокон наркотизированных животных (по Kenshalo в [25] с изменениями) |
|
Рис. 9.13. Реакции холодового рецептора кисти обезьяны на кратковременное снижение температуры кожи. Перепады охлаждения были различной величины, а начальная и конечная температуры всегда 34 °С. Данные для одиночного нервного волокна срединного нерва наркотизированной обезьяны (Darian-Smith. Johnson, Dykes: J. Neurophysiol, 36, 325, 1973) |

цепторов, поскольку даже при температуре ниже максимума статической импульсации охлаждение кратковременно повышает ее частоту. Предполагалось также, что температуры ниже этого максимума распознаются по наличию групповых импульсов (рис. 9.13).
Реакции при изменении температуры кожи. На
рис. 9.13 показаны динамические реакции холодового рецептора во время охлаждения и согревания. Сравнение с рис. 9.12 показывает, что частота импульсации зависит не только от температуры (пропорциональный рецептор), но и от скорости ее изменения (дифференциальный рецептор). Это свойство вытекает и из результатов описанных выше психофизических опытов. Поведение тепловых рецепто-