

Функциональные свойства спинальной соматовисцеральной системы
Соматовисцеральные афференты образуют синаптические связи с нейронами в заднем (дорсальном) роге спинного мозга (рис. 9.19). Некоторые толстые миелинизированные афференты (группы 1 и II) также отделяют ветви (коллатерали), восходящие в составе спинного мозга (см. с. 208). У заднего рога, где идет нейронная переработка информации, четыре выхода: длинные тракты, восходящие до головного мозга главным образом в переднебоковом канатике (см. с. 209); восходящие и нисходящие проприоспинальные связи с соседними сегментами; сегментарные связи с мотонейронами и симпатическими нейронами (участие в спинальных двигательных и вегетативных рефлексах). Однако, по всей вероятности, один нейрон не образует все четыре вида связей сразу.
В восходящих трактах аксоны, идущие от одного сегмента, расположены бок о бок. Это создает
соматотопическую слоистость (рис. 9.19); аксоны, присоединяющиеся к прочим во все более высоких сегментах, всегда оказываются со стороны серого вещества. Однако границы пучков не такие резкие, как показано на рис. 9.19. Один из результатов такого расположения состоит в том, что при поверхностном повреждении переднебокового канатика (раной или опухолью) в шейной области первые сенсорные дефициты следует ожидать в нижней половине тела.
Нейроны переднебокового канатика. Хотя клинические данные (рис. 9.17) указывают на участие переднебокового канатика в болевых и температурных ощущениях, он содержит всего лишь несколько аксонов специфичных ноцицептивных и терморецептнвных нейронов. Большинство аксонов принадлежит здесь клеткам, возбуждаемым также слабыми механическими стимулами, т.е. мультирецептивным нейронам. Остается спорным вопрос, достаточно ли немногих специфичных ноцицептивных и терморецептивных нейронов для передачи соответствующих сигналов или дополнительная информация, требуемая головному мозгу для болевых и температурных ощущений, обеспечивается активностью мультирецептивных нейронов [13, 23, 39].
212 Часть III. Общая и специальная сенсорная физиология
|
Рис. 9.19. Соединения кожных и висцеральных афферентов в спинном мозгу. Входя через задние корешки, афференты образуют синапсы с нейронами заднего рога, которые передают информацию сегментарным спинальным эфферентам, соседним сегментам (по проприоспинальным волокнам) и восходящим трактам (например, переднебокового канатика). Коллатерали аффервнтов групп I и II восходят в составе дорсальных столбов белого вещества непосредственно до продолговатого мозга. Два тормозных влияния на нейрон заднего рога (показаны красным) оказывают нисходящие пути от головного мозга и спинальные интернейроны. Справа схематично показано топографическое разделение восходящих волокон от различных частей спинного мозга в белом веществе его шейного отдела |
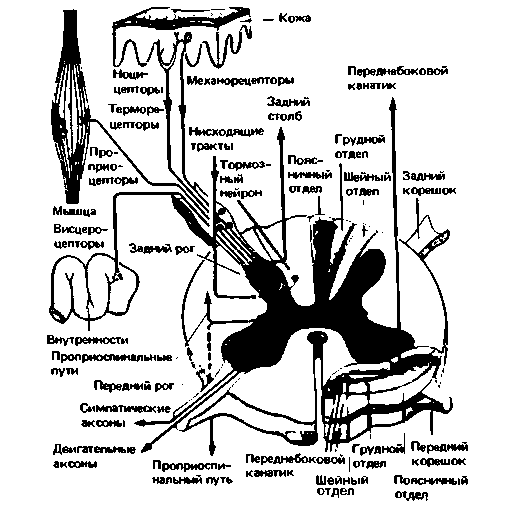
Важное свойство заднего корешка как станции переключения - конвергенция афферентов от кожи и внутренних органов на одни и те же нейроны. Из-за нее афферентные сигналы от внутренностей (например, запускаемые желчными коликами) «ложно» локализуются больным на поверхности тела (отраженная боль, см. гл. 10). Каждый внутренний орган связан с определенной областью кожи — своей зоной Геда. Другой элемент цепи заднего корешка (показан красным на рис. 9.19) - тормозные синапсы, которые служат для модификации афферентной информации. Это может принимать форму латерального торможения (см. гл. 8) или регуляции сенсорного входа трактами, нисходящими от головного мозга (нисходящее торможение, рис. 9.26 и 9.27). Активация таких тормозных механизмов применима для обезболивания (см. гл. 10).
Нейроны ядер заднего столба. Аксоны заднего столба оканчиваются ипсилатерально в его ядрах (клиновидном и тонком), находящихся в продолговатом мозгу. Здесь они образуют синапсы с крупными нейронами, посылающими аксоны в контралатеральный таламус по медиальному лемннсковому тракту (рис. 9.20) и называемыми релейными в отличие от вставочных (интернейронов) с аксонами, не выходящими из ядер заднего столба. Переработка афферентной информации при синаптической передаче к релейным нейронам имеет следующие характеристики:
сохранение сенсорной специфики; на одном нейроне конвергируют афференты одинакового сенсорного типа;
большой запас надежности синаптической передачи; даже одиночные импульсы афферентного волокна вызывают постсинаптическую импульсацию;
мелкие рецептивные поля;
соматотопическая организация; пространственно упорядоченное представительство кожи;
афферентное торможение (латеральное торможение, с. 186);
нисходящие регулирующие входы, в частности, от коры головного мозга (рис. 9.26).
Эти свойства характерны и для последующих мест переключения в лемнисковой системе; они определяют качество и разрешающую способность ощущений, связанных с передаваемой здесь информацией.