

2)Виды мышечных волокон. Иннервация скелетной мышцы. Нейромоторная ( двигательная) единица. Нейротрофический контроль свойств скелетной мышцы.
Виды мышц:
-скелетные: произвольные, поперечно-полосатые
-сердечные: непроизвольные, поперечно-полосатые
-гладкие: непроизвольные, нет поперечной исчерченности
Типы мышечных волокон:
1)Фазные волокна:
- медленные (красные) – много миоглобина ( связывает О2) и митохондрий; поддержание позы; утомление наступает очень медленно, быстро проходит.
- быстрые (окислительного типа) – много митохондрий; образует АТФ путём окислительного фосфорилирования; быстрые энергичные движения без заметного утомления.
- быстрые (гликолитического типа, «белые») – мало митохондрий, миоглобина нет; АТФ образуется за счёт гликолиза; быстрое и сильное сокращение, но сравнительно быстро утомляется.
2) Тонические
- сокращение и расслабление происходят медленно; входят в состав наружных мышц глаза.
Иннервация скелетной мышцы:
Соматическая нервная система (произвольная); один двигательный нейрон (тело в ЦНС); осуществляет возбуждение. Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы - от двигательных нейронов определенных черепных нервов. При этом к каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется одним мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже сотни мышечных волокон получают двигательную иннервацию от одного мотонейрона, посредством разветвления его аксона.
Двигательное нервное волокно, подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обуславливая выход ионов кальция и начало процесса сокращения мышечного волокна.
Нейромоторная единица - совокупность нейрона и группы мышечных волокон, иннервируемых аксоном этого нейрона.
В состав нейромоторной единицы входят:
нервная клетка - в основном мотонейроны, тела которых лежат в передних рогах спинного мозга;
аксон мотонейрона - миелиновые волокна;
группа мышечных волокон - в зависимости от вида деятельности количества волокон различно. Если тонкая работа 2-4, если грубая - до нескольких тысяч.
Виды нейромоторных единиц
По характеру возбуждения, возникающего в мышечных волокнах все нейромоторные единицы делятся на 2 группы.
Фазные нейромоторные единицы- образуются альфа-мотонейронами. Это самые крупные мотонейроны (диаметр 10-20 мкм). Скорость проведения возбуждения по отростку этого аксона - 120 м/с. Аксон обеспечивает одиночную иннервацию, образуя на каждом волокне 1-2 синапса. В каждом синапсе при возбуждении возникает достаточный потенциал концевой пластинки, который обеспечивает возникновение потенциала действия в мышце (импульсное возбуждение). Лучше работают под действием одиночного возбуждения (импульса), при котором возникает распространяющееся возбуждение.
Среди фазных нейромоторных единиц выделяют быстрые и медленные.
Быстрые - продолжительность потенциала действия в 2 раза меньше, чем в медленных. Волна сокращения в 5 раз меньше, чем в медленных. Скорость распространения возбуждения в быстрых нейромоторных единицах в 2 раза больше, чем в медленных. Таким образом, быстрые фазные нейромоторные единицы обеспечивают динамическую работу, когда быстрое сокращение сменяется быстрым расслаблением. Для этого вида нейромоторных единиц характерно анаэробное образование энергии. Эти мышечные волокна практически не содержат миоглобина - светлые, белые мышцы;
Медленные- обеспечивают в основном статическую работу, медленное, длительное сокращение мышц. Основной поставщик энергии окислительно-восстановительные процессы. Содержит миоглобин, который депонирует кислород. По цвету темные, красные мышцы.
Тонические нейромоторные единицы- образуются гамма-мотонейронами передних рогов спинного мозга (самые мелкие, диаметр 4-6 мкм). Скорость проведения импульса по отросткам этих нейронов - 30 м/с, в синапсах возникает местный потенциал концевой пластинки (местное возбуждение). Аксоны тонических нейронов обеспечивают множественную иннервацию, т. е. на каждом мышечном волокне образуются несколько десятков синапсов и за счет суммации местных потенциалов и возникает потенциал действия. Возбуждение тонической нейромоторной единицы происходит под влиянием серии импульсов (частота около 10 Гц). Тонические нейромоторные единицы обеспечивают медленное сокращение мышц, участвуют в возникновении тонуса.
Скелетные мышцы образуются различными мышечными волокнами, которые входят в состав фазных и тонических нейромоторных единиц. Попеременное включение нейромоторных единиц обеспечивает изменение функционального состояния мышцы. В состоянии покоя работают тонические нейромоторные единицы, в состоянии активности - фазные нейромоторные единицы.
Кроме двигательной иннервации присутствует вегетативная. Все скелетные мышцы получают импульсы из симпатической нервной системы, которая регулирует обменные процессы.
Нейротрофический контроль свойств скелетной мышцы.
3)Особенности проведения возбуждения в нервно-мышечном синапсе. Везикулярная гипотеза. Квантовая секреция медиатора. Механизм слияния пресинаптических везикул с постсинаптической мембраной (роль пептидов нервного окончания и мембраны везикул). Спонтанная квантовая секреция.
Синапс – специализированный межклеточный контакт ,обеспечивающий передачу информации между двумя возбудимыми клетками.
Cтруктура синапса:
1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке);
2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс);
3) синаптическая щель (пространство между пресинаптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови).
2. Механизмы передачи возбуждения в синапсах на примере мионеврального синапса (Полный ответ)
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
Проведение возбуждения в НМС ( по лекции)
1.ПД нервного окончания
2.активания потенциал зависимых Са2+ каналов, вход ионов Са2+ в нервное окончание
3.освобождение ацетилхолина путем экзоцитоза везикул в синаптическую щель
4.активания постсинаптических н-холинорецепторов
5.Вход Na+ возникновение потенциала концевой пластинки, затем потенциала действия, вход Са2+
6.сокращение мышцы
Везикулярная гипотеза- 1) нейромедиатор выделяется из нервного окончания мультимолекулярными порчиями – квантами. 2)Один квант находится в одной синаптической везикуле. 3) везикулярный медиатор осовбождается путем слияния мембраны везикулы с пресинаптической мембраной (экзоцитоз)
Кватовая
секреция медиатора
– Са2+ зависимый процесс. Повышение
Са2+ в аксоплазе от
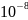 (покой) до
(покой) до
 (активация) вызывает массивное
освобождение квантов медиатора
(экзоцитоз). Основной пусть выхода Са2+
в терминаль – потенциал зависимые Са2+
каналы с пресинаптической мембране,
открываемые при деполяризации. Блокада
этих каналов ( например кадмием) устраняет
эффект стимуляции.
(активация) вызывает массивное
освобождение квантов медиатора
(экзоцитоз). Основной пусть выхода Са2+
в терминаль – потенциал зависимые Са2+
каналы с пресинаптической мембране,
открываемые при деполяризации. Блокада
этих каналов ( например кадмием) устраняет
эффект стимуляции.
Последовательность явлений:
1.Приход ПД в терминаль
2.Активания Са2+ каналов
3.Быстрый локальный рост Са2+
4.Вызванное Са2+ повышение вероятности экзоцитоза медиатора
5.этот процесс занимает 0,2-0,5 мс что соответствует синаптической задержке между стимулом и постсинаптическим ответом
Механизм слияния пресинаптических везикул с постсинаптической мембраной (роль пептидов нервного окончания и мембраны везикул).
В пресинаптическом нервном окончании в синаптических везикулах аккумулируются медиаторы (трансмиттеры). Потенциал действия, достигая нервного окончания, деполяризует его мембрану. Деполяризация вызывает открытие кальциевых каналов (насосов), по которым в окончание входит ток ионов Са2+. Ионы Са2+ вызывают процесс слияния везикул с пресинаптической мембраной с последующим открытием их и выбрасыванием своего содержимого (медиаторов) в синаптическую щель.
Синаптические везикулы фиксируются большей частью на цитоскелете посредством протеина синапсина, локализованного на цитоплазматической поверхности каждой везикулы, к протеину спектрину, расположенному на волокнах F-актина цитоскелета, и образуют тем самым трансмиттерный резервуар. Меньшая часть везикул связана специфическими протеинами с внутренней стороной пресинаптической мембраны. Это взаимодействие осуществляется посредством белка мембраны везикулы синаптобревина и белка пресинаптической мембраны синтаксина . Именно эти везикулы непосредственно поставляют трансмиттер для очередного выброса.
Если потенциал действия достиг пресинаптической области и в пресинаптическом окончании концентрация Са2+ поднялась до необходимого уровня, то происходят два процесса. Во-первых, на уже связанных с пресинаптической мембраной везикулах, которые, по существу, лежат на ней, Са2+ связывается с протеином, входящим в состав их мембраны, - синаптотагмином. Это приводит к тому, что мембрана везикулы раскрывается. Одновременно комплекс полипептида синаптофизина сливается с неидентифицированными протеинами пресинаптической мембраны. При этом возникает пора, через которую осуществляется регулируемый экзоцитоз, т.е. секреция трансмиттера в синаптическую щель, причем еще один протеин везикулы, rab3A , регулирует этот процесс. В одной везикуле сосредоточено примерно 6000-8000 молекул трансмиттера; это наименьшее количество трансмиттера, освобожденного в синаптическую щель, которое называется "один квант трансмиттера ".
В совокупности локальная концентрация трансмиттера в синаптической щели после его освобождения относительно высока и находится в миллимолярном диапазоне.
Таким образом, главную роль для процесса выброса трансмиттера по типу экзоцитоза играет не деполяризация окончания, а именно входящий ток ионов Са2+. Ионы Са2+ служат при этом не для дополнительной деполяризации, а в качестве вещества-посредника (вторичного мессенджера), которое запускает механизм слияния везикул. Повышение концентрации экстрацеллюлярного Са2+ повышает входящий ток ионов Са2+ и, тем самым, увеличивает освобождение трансмиттера. Наоборот, искусственное повышение концентрации экстрацеллюлярного Mg2+ посредством замещения им ионов Са2+ ведет к снижению входящего тока Са2+ и, тем самым, уменьшению освобождения трансмиттера. Небольшие G-белки , вероятно, также участвуют в управлении везикулярным экзоцитозом.
После прекращения пресинаптического потенциала действия ионы Са2+ удаляются из пресинаптической области посредством активного ионного транспорта с участием Са2+-АТФазы и 3Na+/Ca2+-обменника.
Спонтанная квантовая секреция.
Медиатор секретируется мультимолекулярными пакетами-квантами. Квант соответствует содержимому одного синаптического пузырька. МПКП – миниатюрный потенциал концевой пластины – результат постсинаптического действия одного кванта медиатора.
МПКП:
- возникают спонтанно вне зависимости от стимуляции нерва
- имеют тот же временный ход что и вызванные ответы
- амплитула МПКП зависит от состояния АХ-рецепторов и конц. АХ в везикуле
- частота МПКП зависит от состояния терминали
4. На концевой пластинке нервно-мышечного синапса возбуждающий постсинаптический потенциал называют также потенциалом концевой пластинки.В нервно-мышечном синапсе ацетилхолин синтезируется в окончаниях двигательных нервов и накапливается в пузырьках. Когда в окончание приходит потенциал действия, ацетилхолин из 150-200 пузырьков высвобождается в синаптическую щель и связывается с холинорецепторами (холинорецепторы нервно-мышечных синапсов принадлежат к N-холинорецепторам ), плотность которых особенно высока на гребнях складок постсинаптической мембраны. Каналы, сопряженные с холинорецепторами, открываются, в клетку входят катионы (в основном Na+), и происходит деполяризация постсинаптической мембраны, называемая потенциалом концевой пластинки. Поскольку этот потенциал в норме всегда сверхпороговый, он вызывает потенциал действия, распространяющийся по мышечному волокну и вызывающий сокращение. Потенциал концевой пластинки короткий, так как ацетилхолин, во-первых, быстро отсоединяется от рецепторов, во-вторых - гидролизуется АХЭ .Потенциал концевой пластинки аналогичен ВПСП в межнейронных синапсах.Однако амплитуда одиночного ПКП существенно выше, чем ВПСП, потому что в нервно-мышечном соединении высвобождаемый нейромедиатор попадает на более обширную поверхность, где связывается с гораздо большим количеством рецепторов и где, следовательно, открывается намного больше ионных каналов. По этой причине амплитуда одиночного ПКП обычно бывает более чем достаточна для того, чтобы в смежной с концевой пластинкой области плазматической мышечной мембраны возник местный электрический ток, инициирующий потенциал действия. Затем потенциал действия распространяется по поверхности мышечного волокна посредством такого же механизма ( рис. 30.19 ), что и в мембране аксона. Большинство нервно- мышечных соединений расположены в срединной части мышечного волокна, откуда возникший потенциал действия распространяется к обоим его концам.
Миниатюрные потенциалы субсинаптической мембраны - это низкоамплитудные кратковременные изменения полярности субсинаптической мембраны нервной клетки, мышечного волокна в покое. Все характеристики этих миниатюрных затухающих колебательных процессов, в частности моменты времени возникновения, амплитуды, интервалы между МПСМ, являются вероятностными характеристиками. Изменения полярности мембраны нейрона могут быть как в сторону деполяризации , так и в сторону гиперполяризации. В первом случае говорят о деполяризационном МПСМ (ДМПСП). Другое название - возбудительный постсинаптический потенциал, ВПСП. Во втором случае говорят о гиперполяризационном МПСМ (ГМПСП). Другое название - тормозный постсинаптический потенциал, ТПСП. Изменения полярности субсинаптической мембраны мышечной клетки называют миниатюрным потенциалом концевой пластинки.
5. При действии различных факторов на нерв, вызывающих длительную деполяризацию клеточной мембраны, возникает полный блок проведения нервных импульсов. В дальнейшем было показано, что парабиотические состояние связано с инактивацией более 50% натриевых каналов. Для возникновения блока в проведении возбуждения протяженность парабиотического участка должна превысит 5 мм в толстом миелинизированном волокне, иначе ПД может возбуждать соседний участок волокна действием своего электрического поля. Нарушение физиологической непрерывности нервных волокон возникает при действии анестетиков, гипоксии, при воспалении, охлаждении. После действия этих факторов, не вызвавших грубых структурных изменений, проведение возбуждения по волокнам нерва восстанавливается. Это свойство нервных волокон используется в медицинской практике.
ВЕЩЕСТВА, АКТИВИРУЮЩИЕ ПЕРЕДАЧУ ВОЗБУЖДЕНИЯ
Холиномиметики. Метахолин, карбахол и никотин оказывают на мышцу тот же эффект, что и ацетилхолин. Различие заключается в том, что эти вещества не разрушаются ацетилхолинэстеразой или разрушаются более медленно, в течение многих минут и даже часов.
Антихолинэстеразные соединения. Неостигмин, физостигмин и диизопропилфлуорофосфат инактивируют фермент таким образом, что имеющаяся в синапсе ацетилхолинэстераза теряет способность гидролизовать ацетилхолин, выделяющийся в концевой двигательной пластинке. В результате происходит накопление ацетилхолина, что в ряде случаев может вызывать мышечный спазм. Это может приводить к смертельным исходам при спазме гортани у курильщиков. Неостигмин и физостигмин инактивируют ацетилхолинэстеразу в течение нескольких часов, после чего их действие проходит, и синаптическая ацетилхолинэстераза восстанавливает свою активность. Диизопропилфлуорофосфат, являющийся нервно-паралитическим газом, блокирует ацетилхолинэстеразу на недели, что делает это вещество смертельно опасным.
ВЕЩЕСТВА, БЛОКИРУЮЩИЕ ПЕРЕДАЧУ ВОЗБУЖДЕНИЯ
Миорелаксанты периферического действия (кураре и курареподобные препараты) широко применяются в анестезиологии. Тубокурарин препятствует деполяризующему действию ацетилхолина. Дитилин приводит к миопаралитическому эффекту, вызывая стойкую деполяризацию постсинаптической мембраны.Ботулотоксин и столбнячный токсин блокируют секрецию медиатора из нервных терминалей.Бетта- и гамма-Бунгаротоксины блокируют холинорецепторы.
6. В зависимости от сократительных свойств, гистохимической окраски и утомляемости мышечные волокна подразделяют на две группы - красные и белые. Все мышечные волокна двигательной единицы принадлежат к одному типу.Функциональной единицей мышечного волокна является миофибрилла . Миофибриллы занимают практически всю цитоплазму мышечного волокна, оттесняя ядра на периферию
Различают два типа мышечных волокон.
Красные мышечные волокна (волокна 1 типа) содержат большое количество митохондрий с высокой активностью окислительных ферментов. Сила их сокращений сравнительно невелика, а скорость потребления энергии такова, что им вполне хватает аэробного метаболизма. Они участвуют в движениях, не требующих значительных усилий, - например, в поддержании позы.
Белым мышечным волокнам (волокнам 2 типа) присуща высокая активность ферментов гликолиза, значительная сила сокращения и такая высокая скорость потребления энергии, для которой уже не хватает аэробного метаболизма. Поэтому двигательные единицы , состоящие из белых волокон, обеспечивают быстрые, но кратковременные движения, требующие рывковых усилий.
Плавные произвольные движения начинаются с активации красных волокон. Если (как в норме) окончание мотонейрона выделяет достаточное количество ацетилхолина , а на постсинаптической мембране имеется необходимое количество холинорецепторов , происходит пороговая деполяризация постсинаптической мембраны и возникает потенциал действия. Последний распространяется по мембране мышечного волокна, затем по поперечным трубочкам переходит внутрь и запускает процессы электромеханического сопряжения, заканчивающиеся сокращением мышечного волокна.
Каждая миофибрилла имеет периодическое строение. Повторяющаяся структура в составе миофибриллы называется саркомером . Саркомеры соседних миофибрилл расположены друг против друга, отчего все мышечное волокно тоже приобретает периодическое строение.
Функции мышц регулируются различными отделами центральной нервной системы (ЦНС), которые во многом определяют характер их разносторонней активности (фазы движения, тонического напряжения и р.). Рецепторы двигательного аппарата дают начало афферентным волокнам двигательного анализатора, которые составляют 30-50% волокон смешанных (афферентно-эфферентных) нервов, направляющихся в спинной мозг. Сокращение мышц вызывает импульсы, которые являются источником мышечного чувства - кинестезии.
Передача возбуждения с нервного волокна на мышечное осуществляется через нервно-мышечный синапс ,который состоит из двух разделенных щелью мембран - пресинаптической (нервного происхождения) и постсинаптической (мышечного происхождения). При воздействии нервного импульса выделяются кванты ацетилхолина, который приводит к возникновению электрического потенциала, способного возбудить мышечное волокно. Скорость проведения нервного импульса через синапс в тысячи раз меньше, чем в нервном волокне. Он проводит возбуждение только в направлении к мышце. В норме через нервно-мышечный синапс млекопитающих может пройти до 150 импульсов в одну секунду. При утомлении (или патологии) подвижность нервно-мышечных окончаний снижается, а характер импульсов может изменяться.
«Теория скользящих нитей» - концепция, объясняющая механизм сокращения миофибриллы. Разработана в 1954 г. независимо друг от друга Хью Эзмором Хаксли и Сэром Андру Филдингом Хаксли (Н. Е. Huxley, 1924- , английский молекулярный биолог, Sir А. F. Huxley, 1917- , английский физиолог, лауреат Нобелевской премии по физиологии и медицине 1963 г). Согласно данной концепции, укорочение саркомера во время сокращения происходит благодаря активному скольжению актиновых нитей относительно миозиновых нитей. Укорочение заканчивается, когда актиновые нити втягиваются к центру саркомера. При расслаблении или растяжении мышцы область взаимного перекрывания тонких и толстых филаментов пассивно суживается. В настоящее время существуют модели механизма управления взаимодействием актиновых и миозиновых нитей, обеспечивающих сокращение миофибриллы, однако некоторые аспекты этих гипотез остаются пока недоказанными.
7. Решающую роль в мышечном сокращении играют ионы кальция. В мышечном волокне, кроме саркоплазматического ретикулума, имеются поперечные каналы, возникшие из углублений сарколеммы. Активность АТФ-азы регулируется изменением концентрации ионов кальция. В стадии покоя ионы кальция накапливаются в саркоплазматическом ретикулуме. При сокращении они перемещаются в волокна миозина, вызывая активацию АТФ-азы. В результате этого происходит сокращение; затем ионы кальция поступают в саркоплазматический ретикулум, чтобы начать новый цикл мышечного сокращения. Итак, под влиянием ионов кальция укрепляются связи между субъединицами тропонина и одновременно ослабляется контакт тропонина с актином, поэтому последний может занять «открытое» положение. В свою очередь при снижении концентрации ионов кальция взаимодействие субъединиц тропонина ослабляется. Это позволяет субъединице Tnl вступить с актином в прочную связь, вызывая смещение тропомиозина и переход актина в «закрытое» положение, когда реагирова ние с головками миозина невозможно. Роль тропомиозина в этом процессе заключается в передаче блокирующего или деблокирующего эффекта тропонина одновременно семи молекулам актина. В гладкой мышце, как и в скелетной, сократительный процесс активируется ионами кальция, однако источники этих ионов различны. Различие заключается в том, что саркоплазматический ретикулум, обеспечивающий практически все количество ионов кальция для сокращения скелетной мышцы, в большинстве гладких мышц очень слабо развит. В гладкой мышце, как и в скелетной, сократительный процесс активируется ионами кальция, однако источники этих ионов различны. Различие заключается в том, что саркоплазматический ретикулум, обеспечивающий практически все количество ионов кальция для сокращения скелетной мышцы, в большинстве гладких мышц очень слабо развит. Вместо этого почти все ионы кальция, вызывающие сокращение, входят в мышечную клетку из внеклеточной жидкости во время потенциала действия или под влиянием другого стимула. Электромеханическое сопряжение - это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску цикла поперечных мостиков .Электрическая активность плазматической мембраны не оказывает прямого влияния на сократительные белки, а вызывает повышение цитоплазматической концентрации ионов Са2+.
8 Единственным непосредственным источником энергии для мышечного сокращения служит АТФ. Запасов АТФ в мышцах очень мало — хватает на поддержание их работы в течение долей секунды. Поэтому для обеспечения длительной деятельности мышц необходимо постоянное восстановление АТФ. Этот процесс осуществляется в мышцах анаэробным (без участия кислорода) и аэробным (с участием кислорода) путями. В организме человека могут использоваться три основных источника «топлива»: креатин-фосфат (КрФ), углеводы в виде гликогена и глюкозы и жиры. Энергетических систем получения АТФ в работающей мышце тоже три: фосфогенная, гликоли-тическая и окислительная. Восстановление АТФ во время мышечного сокращения происходит почти мгновенно за счет КрФ. При этом работает фосфогенная система, которая способна осуществить восстановление АТФ со скоростью, необходимой для выполнения работы, которая характеризуется максимальной мощностью Обеспечения мышц энергией при более продолжительной физической нагрузке поддерживается за счет гликолитической системы. В основе ее деятельности лежит процесс анаэробного расщепления углеводов (гликогена и глюкозы) до молочной кислоты. При увеличении продолжительности работы энергообеспечение мышц в основном осуществляется за счет окислительной системы и процесса, называемого окислительным фосфорилированием. Это аэробный процесс, то есть он осуществляется при достаточном снабжении мышц кислородом. В качестве энергетических субстратов используются углеводы. Удаление ионов Са2+ от сократительных белков приводит к началу расслабления. При расслаблении концентрация кальция в саркоплазме снижается от 10-5 до 10-7моль*л-1. Это приводит к потере активности АТФ-азы миозина. Головки миозиновых нитей связывают АТФ, но не расщепляют их. Тропонин при отсутствии кальция снова блокирует активные центры тонких нитей. Все это приводит к разрыву актомиозиновых комплексов и расхождению актиновых и миозиновых нитей. Под действием упругих сил белков стромы мышца возвращается в исходное состояние. Таким образом, в процессе сокращения и расслабления мышц АТФ выполняет следующую роль: - в покоящейся мышце – препятствует соединению актиновых нитей с миозиновыми; - в процесс сокращения мышцы – поставляет необходимую энергию для движения тонких нитей относительно толстых, что приводит к укорочению мышцы или развитию напряжения; Процесс мышечного расслабления В настоящее время расслабление считается активным процессом, причем расход энергии больше, чем при сокращении. Источник сокращения - АТФ, расщепляющийся при сокращении. Трупным окоченением принято называть состояние мышц трупа при котором они уплотняются и фиксируют части трупа в определенном положении. Окоченевшее мертвое тело как бы деревенеет. Непосредственно после наступления смерти все мышцы тела человека расслабляются, теряют свойственную им прижизненную упругость, лицо принимает спокойный вид, отсюда, наверное, происходит слово покойник. После прекращения основных процессов жизнедеятельности во всех мышцах тела начинаются сложные биохимические процессы, связанные с преобразованием аденазинтрифосфорной кислоты (АТФ) и высокомолекулярных структур актина и миозина.
9 Механический ответ отдельного мышечного волокна на одиночный потенциал действия называется одиночным сокращением. Последовательность явлений во время изотонического одиночного сокращения. При возбуждении мышечного волокна поперечные мостики начинают развивать силу, однако укорочение не начнется, пока мышечное напряжение не превысит нагрузку на волокно. Таким образом, укорочению предшествует период изометрического сокращения , в течение которого возрастает напряжение. Суммация силы. Суммация означает сложение отдельных одиночных сокращений, ведущее к увеличению интенсивности общего сокращения мышцы. Суммация осуществляется двумя путями: (1) путем увеличения числа моторных единиц, сокращающихся одновременно, что называют суммацией сокращений многих волокон; (2) путем увеличения частоты сокращений, что называют временной (частотной) суммацией, которая может привести к тетанизации. Тетанус, тетаническое мышечное сокращение— состояние длительного сокращения, непрерывного напряжения мышцы, возникающее при поступлении к ней через мотонейрон нервных импульсов с высокой частотой. При этом расслабления между последовательными одиночными сокращениями не происходит и возникает их суммация, приводящая к стойкому максимальному сокращению мышцы. Различают зубчатый и гладкий тетанус. При зубчатом тетанусе каждый последующий нервный импульс воздействует на начавшую расслабляться мышцу, при этом происходит неполная суммация сокращений. При гладком тетанусе, имеющем бо́льшую амплитуду, воздействие импульса происходит в конце периода укорочения, что приводит к полной суммации сокращений. Пессимальное торможение Деполяризация постсинаптической мембраны при очень частом следовании друг за другом нервных импульсов лежит в основе открытого Н. Е. Введенским пессимального торможения. Это явление часто называют торможением Введенского. Сущность его состоит в следующем. Величина тетанического сокращения скелетной мышцы в ответ на ритмические раздражения нерва возрастает с увеличением частоты стимуляции. При некоторой оптимальной частоте раздражения тетанус достигает наибольшей величины. Если продолжать увеличивать частоту стимуляции нерва, то тетаническое сокращение мышцы начинает резко ослабевать и при некоторой большой пессимальной частоте раздражения нерва мышца, несмотря на продолжающееся раздражение, почти пол ностью расслабляется. Уменьшение частоты стимуляции тотчас приводит к восстановле нию высокого уровня тетанического сокращения.