

Особенности электромеханического сопряжение при сокращении сердечного миоцита
У миокардиоцита значительное колличество Ca2+ в цитозроль поступает из интерстиция (рис. 709270937).
Рис. 709270937. Пути поступления Ca2+ в цитозоль у разных типов миоцитов при сокращении.
|

Рианодиновые рецепторы, расположенные на мембране саркоплазматического ретикулума, активируются входящим из межклеточного вещества Ca2+.a
Кофеин также оказывает на эти рецепторы активирующее действие.
Особенности электромеханического сопряжение при сокращении гладкого миоцита
У гладких миоцитов основное колличество Ca2+ в цитозроль поступает из интерстиция (рис. 709270937).
Каналы Ca2+ на мембране плохо выраженного саркоплазматического ретикулума управляются рецепторами инозитолтрифосфата b (ИТФ). При возбуждении плазматической мембраны активируется фосфолипаза С и образуется ИТФ.
Этот механизм функционирует в лимфоцитах, овоцитах.
10. Рабочий цикл миозиновых (поперечных) мостиков
|
16 |
|
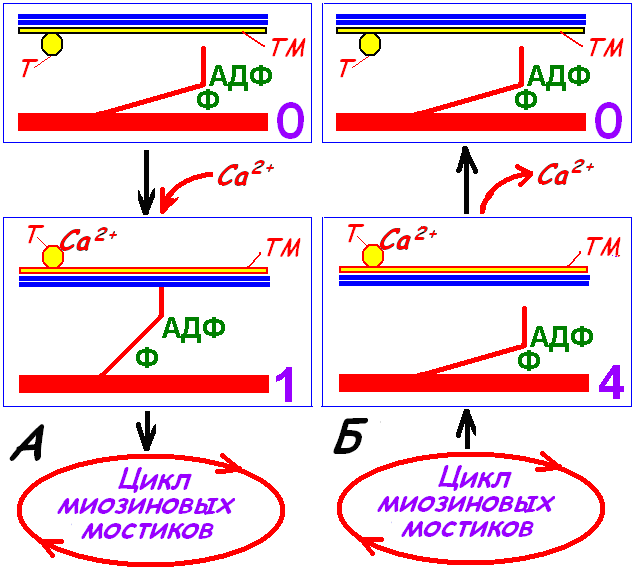
Вхождение актомиозинового комплекса в цикл миозиновых головок показан на рис. 709270916 А.
Рис. 709270916. Вхождение актомиозинового комплекса в цикл миозиновых головок (А) и выход из цикла (Б) поперечнополосатого миоцита.
|
.
Рабочий цикл миозиновых мостиков поперечнополосатого миоцита
Рабочий цикл миозиновых мостиков поперечнополосатого миоцита включает следующие процессы (рис. 709270802):
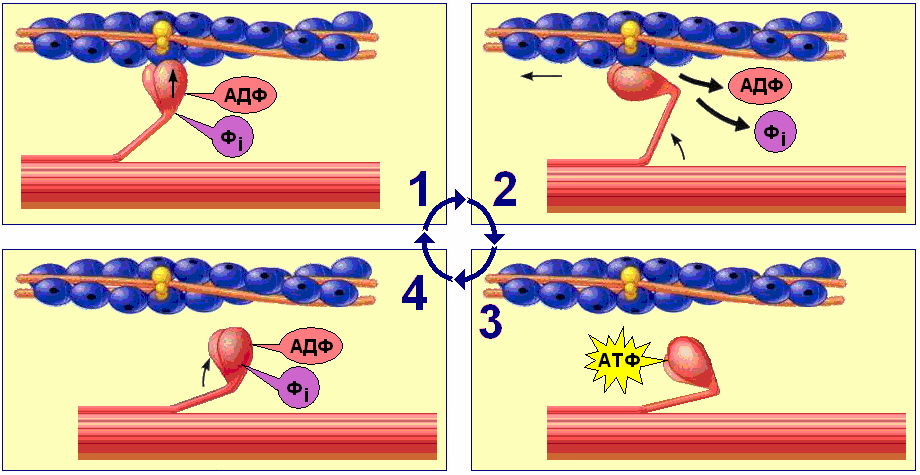
Рис. 709270802. Рабочий цикл миозиновых мостиков поперечнополосатого миоцита.
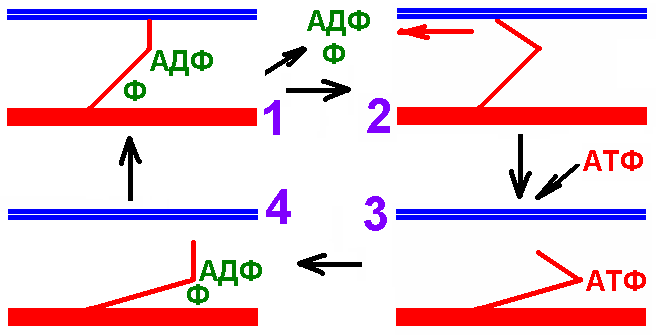
Рис. 709270834 показывает цикл миозиновых мостиков без деталей и может быть использован студентом при ответе.
Рис. 709270834. Рабочий цикл миозиновых мостиков поперечнополосатого миоцита.
|
1. Головка миозина, несущая продукты гидролиза АТФ (АДФ + фосфат) соединяется с миозинсвязывающим участкам актиновой нити. 2. АДФ и фосфат покидают миозиновую головку. Головка миозина изменяет конформацию за счёт шарнирного участка в области шейки миозина. Происходит гребковое движение, продвигающее актиновую нить к центру саркомера (рабочий ход). 3. Головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. 4. Гидролиз АТФ восстанавливает конформацию молекулы миозина, и она оказывается готовой вступить в новый цикл.
|
Головка миозина совершает около пяти циклов в секунду.
Когда одни головки миозина толстой нити производят тянущее усилие, другие в это время свободны и готовы вступить в очередной цикл.
Следующие друг за другом гребковые движения стягивают тонкие нити к центру саркомера. Скользящие тонкие нити тянут за собой Z-линии, вызывая сокращение саркомера. Поскольку в процесс сокращения практически одномоментно вовлечены все саркомеры мышечного волокна, происходит его укорочение.
До тех пор, пока в межфибриллярном пространстве имеется Ca2+, циклы будут повторяться. Это быстрое прикрепление, разрушение и новое прикрепление поперечных мостиков происходит повторно с большой скоростью и во многих участках миофиламентов во время мышечного сокращения и прекращается, когда Ca2+ уходит из межфибриллярного пространства в цистерны или когда молекулы АТФ образуются не столь быстро, чтобы полностью обеспечивать «зарядку» энергией поперечные мостики.
Напряжение, развиваемое мышечным волокном, зависит от числа замкнутых поперечных мостиков, а скорость развития напряжения — от числа замкнутых поперечных мостиков, образуемых в единицу времени, т.е. от скорости их прикрепления к актину. По-видимому, даже при максимальном возбуждении только часть поперечных мостиков эффективно связана с актиновыми миофиламентами. При повышении скорости укорочения мышцы число прикрепленных поперечных мостиков в каждый момент времени уменьшается. Это и объясняет падение силы сокращения мышцы с увеличением скорости ее укорочения.
Если же мышца не укорачивается (при изометрическом сокращении), напряжение в мышечных волокнах создается за счет повторного прикрепления поперечных мостиков на одних и тех же фиксированных участках актиновых нитей. В этом случае никакой внешней работы не совершается, а энергия (АТФ) расходуется на поддержание напряжения мышцы. Тяга, развиваемая сократительными элементами в миофибриллах, передается сарколемме и через нее — на сухожилия мышц.
Период химико-механических изменений в сократительном миофибриллярном аппарате мышечного волокна, на протяжении которого в нем возникает и поддерживается активная механическая тяга, определяющая процесс сокращения, называется периодом активного состояния.