

Нервная система
Часть 1. Нервная ткань. Общая характеристика и морфо-функциональная характеристика нейронов
2. Межнейронные связи. Синапсы
3. Секреторные нейроны
4. Глия
5. Нервные волокна и окончания
6. Гистогенез нервной ткани (нейро- и глиогенез)
1. Общая характеристика и морфофункциональная характеристика нейронов
Чрезвычайно разнообразные по строению и функции нервные клетки составляют основу центральной (головной и спинной мозг) и периферической нервной систем. Совместно с нейронами при описании нервной ткани рассматриваются второй ее важный компонент – глиальные клетки. Они подразделяются на клетки макроглии – астроциты, олигодендроциты, эпендимоциты и клетки микроглии.
Основные функции нервной системы, осуществляемые нейронами – возбуждение, его проведение и передача импульсов на эффекторные органы Нейроглиальные клетки способствуют выполнению нейронами этих функций. Деятельность нервной системы основана на принципе функционирования рефлекторной дуги, состоящей из нейронов, связанных друг с другом посредством специализированных контактов – синапсов различного вида.
Нейроны позвоночных и большинства беспозвоночных животных, как правило, клетки с многими длинными, сложно ветвящимися отростками, часть которых воспринимает возбуждение. Они называются дендритами, а один из отростков, отличающийся большой длиной и разветвлениями в терминальных отделах, именуется аксоном (рис. 9.1).
Основные функциональные свойства нейронов связаны с особенностью строения их плазматической мембраны, содержащей огромное число потенциал- и лигандзависимых рецепторных комплексов и ионных каналов, а также со способностью выделять в определенных участках (синапсах) нейромедиаторы и нейромодуляторы. Познание структурной организации нервной ткани во многом было обусловлено применением специальных методов окраски нейронов и глиальных клеток. Среди них особого внимания заслуживают методы импрегнации тканей солями серебра по Гольджи и Бильшовскому-Гроссу.
Основы классических представлений о клеточном устройстве нервной системы были заложены в трудах выдающегося испанского нейрогистолога, лауреата Нобелевской премии, Сантьяго Рамон-и-Кахала. Большой вклад в учение о нервной ткани внесли исследования гистологов Казанской и Петербургской-Ленинградской школ нейрогистологии – К. А. Арнштейна, А. С. Догеля, А. Е. Смирнова, Д. А. Тимофеева, А. Н. Миславского, Б. И. Лаврентьева, Н. Г. Колосова, А.А. Заварзина, П.Д.Дейнеки, Н.В. Немилова, Ю.И. Орлова, В.П. Бабминдры и др.
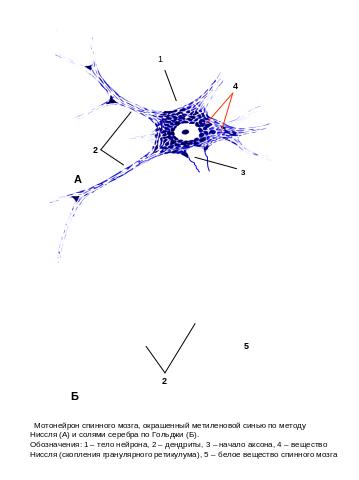
Структурная и функциональная полярность большинства нервных клеток обусловила традиционное выделение трех отделов нейрона: тела, дендритов и аксона (рис. 9.5). Уникальность строения нейронов проявляется в чрезвычайной разветвленности их отростков, нередко достигающих очень большой длины, и наличием в клетках разнообразных специфических белковых и небелковых молекул (нейромедиаторы, нейромодуляторы, нейропептиды и др.), обладающих высокой биологической активностью.
В основе классификации нервных клеток по их строению лежат: 1) форма тела – выделяют округло-овальные, пирамидные, корзинчатые, веретеновидные, грушевидные, звездчатые и некоторые другие виды клеток (рис. 9.5); 2) число отростков – униполярные, биполярные (как вариант – псевдоуниполярные), и мультиполярные; 3) характер ветвления дендритов и наличие шипиков (густо- и редковетвистые; шипиковые и бесшипиковые клетки); 4) характер ветвления аксона (ветвление только в терминальной части или наличие коллатералей по всей длине, короткоаксонные или длинноаксонные) (рис. 9.6).
Нейроны также подразделяют по содержанию нейромедиаторов на: холинергические, адренергические, серотонинергические, ГАМК (гаммкергические), аминокислотные (глицинергические, глутаматэргические и др.). Наличие в одном нейроне нескольких нейромедиаторов, даже таких антагонистических по своим эффектам, как ацетилхолин и норадреналин, заставляет относиться к однозначному определению нейромедиаторного и нейропептидного фенотипа нейронов весьма осторожно.
Также существует классическое разделение нейронов (в зависимости от их положения в рефлекторной дуге) на: афферентные (чувствительные), вставочные (ассоциативные) и эфферентные (в том числе и двигательные). Чувствительные нейроны имеют наиболее вариабельную структурную организацию окончаний дендритов, принципиально отличающую их от дендритов остальных нервных клеток. Они часто представлены биполярными (чувствительные ганглии ряда органов чувств), псевдоуниполярными (спинномозговые ганглии) или высокоспециализированными нейросенсорными клетками (фоторецепторы сетчатки или обонятельные клетки). Найдены нейроны центральной нервной системы, не генерирующие потенциал действия (бесспайковые нейроны), и спонтанно-возбудимые осцилляторные клетки. Анализ особенностей их структурной организации и взаимосвязи с «традиционными» нейронами является перспективным направлением в познании деятельности нервной системы.
Тело (сома). Тела нервных клеток могут значительно различаться по форме и размерам. Моторные нейроны передних рогов спинного мозга и гигантские пирамиды коры больших полушарий – одни из самых крупных клеток в организме позвоночных – размер тела пирамид достигает 130 мкм, и наоборот, клетки-зерна мозжечка, имеющие диаметр в среднем 5–7 мкм, самые маленькие нервные клетки позвоночных. Разнообразны по форме и размерам и клетки вегетативной нервной системы (рис. 9.7).
Ядро. Нейроны имеют, как правило, одно ядро. Оно обычно крупное, округлое, содержит одно-два ядрышка, хроматин отличается низкой степенью конденсации, что свидетельствует о высокой активности ядра (рис. 9.5). Возможно, что некоторые нейроны являются полиплоидными клетками. Ядерная оболочка представлена двумя мембранами, разделенными перинуклеарным пространством и имеющие многочисленные поры. Количество пор достигает у нейронов позвоночных 4000 на ядро. Важной состовляющей ядра является т.н. «ядерный матрикс» - комплекс ядерных белков, обеспечивающих структурную организацию всех компонентов ядра и участствующих в регуляции процессов репликации, транскрипции и процессинге РНК и их выведении из ядра.
Цитоплазма (перикарион). Многие, особенно крупные пирамидные нейроны, отличаются богатым содержанием гранулярной эндоплазматической сети (ГЭС). Это находит яркое проявление при их окраске анилиновыми красителями в виде базофилии цитоплазмы и включенном в нее базофильным, или тигроидным, веществом (вещество Ниссля). Распределение базофильного вещества Ниссля в цитоплазме перикариона признается одним из критериев дифференцировки нейрона, а также показателем функционального состояния клетки. В нейронах находится также большое число свободных рибосом, обычно собранных в розетки – полисомы. В целом, нервные клетки содержат все основные органеллы, характерные для эукариотической животной клетки (рис. 9.8, а), хотя есть ряд особенностей.
Первая касается митохондрий. Интенсивная работа нейрона связана с большими энергетическими затратами, поэтому в них много митохондрий самого разного вида. В теле и отростках нейронов располагаются немногочисленные (3-4 шт) гигантские митохондрии «ретикулярного» и «нитчатого» типов. Расположение крист в них продольное, что также достаточно редко встречается среди митохондрий. Кроме того, в теле и отростках нейрона есть множество мелких митохондрий «традиционного» типа с поперечными кристами. Особенно много митохондрий скапливается в районах синапсов, узлов ветвления дендритов, в начальном участке аксона (аксоном холмике). Из-за интенсивности функционирования митохондрий в нейроне они имеют, как правило, короткий жизненный цикл (некоторые митохондрии живут около часа). Обновляются митохондрии путем традиционного деления или почкования митохондрий и поставляются в отростки клетки посредством аксонального или дендритного транспорта.
Еще одной из характерных черт строения цитоплазмы нейронов позвоночных и беспозвоночных животных является присутствие внутриклеточного пигмента – липофусцина. Липофусцин относится к группе внутриклеточных пигментов, главным составляющим которых являются каротиноид желтого или коричневого цвета. Он находится в мелких мембранозных гранулах, рассеянных по цитоплазме нейрона. Значение липофусцина активно обсуждается. Считается, что это пигмент «старения» нейрона и связан он с процессами неполного расщепления веществ в лисосомах.
В процессе жизненного цикла нервных клеток количество липофусциновых гранул достоверно увеличивается и по их распределению в цитоплазме можно косвенно судить о возрасте нейрона.
Выделяют четыре морфологические стадии «старения» нейрона. У молодых нейронов (1- я стадия - диффузная) - липофусцина мало и он рассеян по цитоплазме нейрона. У зрелых нервных клеток (2-я стадия, околоядерная) - количество пигмента увеличивается и он начинает скапливаться в зоне ядра. У стареющих нейронов (3-я стадия - полярная), липофусцина все больше и больше и скопления его гранул концентрируются около одного из полюсов нейрона. И наконец, у старых нейронов (4-я стадия, биполярная), липофусцин заполняет большой объем цитоплазмы и его скопления находятся на противоположных полюсах нейрона. В ряде случаев липофусцина в клетке становится так много, что его гранулы деформируют ядро. Накопление липофусцина в процессе старения нейронов и организма связывают также со свойством липофусцина, как каротиноида, связывать кислород. Полагают, что таким образом нервная система адаптируется к происходящему с возрастом ухудшению кислородного питания клеток.
Особой разновидностью эндоплазматической сети, характерной для перикариона нейронов, являются субповерхностные цистерны – одна-две уплощенные мембранные везикулы, расположенные около плазматической мембраны и нередко связанные с ней электронно-плотным неоформленным материалом (рис. 9.8, б). В перикарионе и в отростках (аксоне и дендритах) нередко обнаруживаются мультивезикулярные и мультиламеллярные мембранозные тельца, представленные скоплениями пузырьков или фибриллярного материала со средним диаметром 0,5 мкм. Они являются производными конечных стадий функционирования лизосом в процессах физиологической регенерации компонентов нейрона и участвуют в обратном (ретроградном) транспорте. (рис. 9.9).
Дендриты. Дендриты многих нейронов отличаются высокой степенью разветвленности и сложностью окончаний и характеризуются постепенным уменьшением диаметра претерминальных веточек. Дендритная система определенных типов нервных клеток, как правило, более стабильна, чем их аксонная система. Это послужило основой для создания современных классификаций нейронов (классификации Г.И. Полякова, Т.А.Леонтович и Э. Рамон-Молинера). Цитоплазма дендритов во многом аналогична таковой тела клетки и содержит многие органеллы (рис. 1), в том числе гранулярную эндоплазматическую сеть, комплекс
Гольджи, особенно в крупных первичных веточках и узлах ветвления. Особенности структурной организации дендритов проявляются и на молекулярном уровне – в опытах с использованием метода «гибридизации in situ» было показано, что большинство цитоплазматических мРНК определялось в начальных отделах крупных дендритов и только несколько видов мРНК были обнаружены в их маленьких веточках. Эти мРНК отвечают за синтез высокомолекулярной формы МАР2 – белка (микротрубочко - ассоциированный белок, считающийся маркером дендритов) и синтез -субъединицы Ca2+/кальмодулинзависимой протеинкиназы. В дендритах также выявлена мозговая цитоплазматическая РНК (BC1), впервые обнаруженная в мозге крыс и позже показанная у человека под названием BC200. Эта очень короткая РНК (всего 152 нуклеотида) отличается тем, что не кодирует какого-либо белка. Значение этой РНК в дендритах окончательно не установлено. Предполагается, что она может участвовать в регуляции транспорта мРНК в теле и отростках нейронов. Интересно, что нетранслируемые дендритные РНК связаны с белками и образуют рибонуклеопротеидные комплексы, аналогичные частичкам, распознающим сигнальный пептид при синтезе секреторного белка.
Дендриты многих нейронов имеют маленькие цитоплазматические выросты – шипики (по англ. spine – вырост). Количество шипиков на дендритах у разных типов клеток может колебаться от нуля (например у т.н. «безшипиковых короткоаксонных звездчатых нейронов неокортекса высших млекопитающих и человека) до нескольких десятков тысяч (например на дендритах клеток Пуркинье коры мозжечка или крупных радиальных нейронах подкорковых центров полушарий).
Морфологически шипики имеют разнообразную форму и на светооптическом уровне выделяют несколько типов шипиков (палочковидный, грибовидный, конусовидный и другие типы). Объемная,реконструкция шипиков показала, что их размеры и объем значительно варьируют от 0,001 мкм3 у мелких тонких шипиковидных выростов до 0,8 мкм3 у крупных грибовидных шипиков).
Наиболее часто встречаемой формой шипика является грибовидный шипик. Он имеет головку и шейку. Электронно-микроскопические исследования и данные иммуногистохимии выявили сложную внутреннюю структуру шипиков (Рис. хх ).
В большинстве случаев шипики являются постсинаптической частью аксон-дендритного контакта (на каждом шипике может быть от одного до нескольких
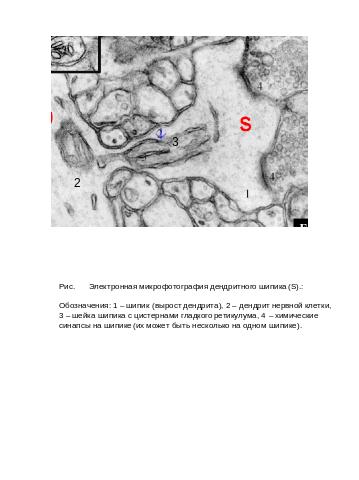
аксонных окончаний). Характерной структурой шипика является развитая система постсинаптических утолщений (PSD). Она занимает около 10% поверхности шипика и имеет сложную молекулярную структуру. Основными компонентами PSD являются встроенные в мембрану постсинапса рецепторные комплексы, ионные каналы, молекулы межклеточной адгезии, а также система субмембранного цитоскелета, стабилизирующего эти рецепторы в мембране и осуществляющие связь с компонентами цитоскелета самого шипика.
В головке большинства шипиков (особенно, крупных грибовидных) обнаружены элементы гладкого эндоплазматического ретикулума – т.н. «шипиковый аппарат - ША». Он представляет собой систему уплощенных цистерн, разделенных прослойкой электронноплотного вещетва. Наличие в шипиках ША является показателем высокого уровня дифференцировки дендритной системы нейронов. Встречается ША преимущественно в дендритных шипиках нейронов высших позвоночных и беспозвоночных животных. В головке шипика также обнаруживаются отдельные мембранные пузырьки, мультивезикулярные тельца и рибосомы. Последнее очень важно, т.к.показывает на возможность локального синтеза белков непосредственно в шипике. Митохондрии в шипиках редки, но их присутствие обычно в период роста шипиков и формирования синаптических контактов.
Цитоскелет шипика очень сложный. Основу его составляет актиновый скелет, включающий два основых компонента. Меньшая (стабильная) часть актинового цитоскелета связана с такими актинассоциированными белками как: спектрин, актинин и др. Именно она обеспечивает стабильность формы шипика, тогда как другая (большая) часть актиновых фибрилл может в течение короткого времени (нескольких минут) реорганизовываться, обеспечивая лабильность и изменчивость шипиков. Эта динамическая популяция актиновых филаментов регулируется через систему актинсвязывающих белков (гельзолин, профилин, кофилин и др.). Большинство этих белков являются Са+2 зависимыми, а депо ионов Са находится в цистернах ША. В ножке шипика и прилегающем к ней районе дендрита обнаружен миозин(стенин), взаимодействие которого с актином способствует изменению размера шипика, и как следствие, изменению ширины синапстической щели аксо-шпикового контакта. Лабильность цитоскелета шипика также связана с процессами формирования шипиков или их редукции.
На формирование шипиков, их редукцию или образование новых влияет множество факторов, таких как: возраст организма, синаптическая активность, информационная нагрузка на нейронные цепи, гормональный фон, температура и многие другие.

Значительное влияние на структуру шипиков оказывают и патологические процессы (особенно нейродегенеративные заболевания наследственной природы), а также токсикологические факторы (наркотики, алкоголь, яды различной природы).
Наиболее типичными проявлениями патологических изменений в морфологии шипиков являются:
- уменьшение количества шипиков на дендритных ветвях, вплоть до их полной редукции (алкоголизм, деафферентация, нейродегенеративные заболевания типа болезни Альцгеймера, эпилепсия и др.)
- уменьшение размеров шипиков (синдром Дауна, некоторые формы шизофрении, сенсорная депривация)
- патологическое, уродливое изменение формы шипиков (алкоголизм, эпилепсия, прионовые болезни, нарушение кровоснабжения мозга (инсульт), голодание)
- слияние шипиков в конгломераты, варикозности (ушибы и черепно-мозговые травмы, острые отравления, гипоксия/ишемия мозга). (Рис. ХХ К Харрис, 2002, BRR 39.
Во всех этих случаях серьезные изменения происходят и во внутренней структуре шипиков: в головке шипиков гипертрофируются цистерны шипикового аппарата, накапливаются мультивезикулярные тела (являющиеся показателем деструктивных процессов в цитоплазме), резко повышается электронная плотность цитоплазмы шипика или, наоборот, происходит отек головки шипика.
Все эти морфологические изменения, без сомнения, сказываются на функции нейронных центров, поскольку нарушается вся система связи между нейронами и отделами мозга.
Формирование шипиков (спиногенез)
В норме в головном мозге происходит постоянный, и довольно интенсивный, процесс редукции шипиков и образования новых, что свидетельствует о больших пластических возможностях нервной ткани.
Имеется несколько путей формирования дендритных шипиков.
1 Развитие шипиков из тонких выростов дендритов (филлоподий). Во время ранних этапов синаптогенеза в развивающемся мозге дендриты образуют многочисленные тончайшие выросты – филлоподии, которые в течение нескольких минут вырастают и пропадают. При формировании синапсов, количество

филлоподий резко уменьшается, тогда как количество стабильных зрелых шипиков увеличивается, что подтверждает роль этих тонких выростов как предшественников пшипиков. Подобные филлоподии обнаруживаются и во взрослом мозге. Функции высокоподвижных филлоподий заключаются, возможно, в мобильном поиске в окружающем дендрит пространстве мест контакта с проходящими аксонами и формирование новых синаптических контактов.
2. Развитие шипиков из дендритного ствола. В этом случае шипик развивается в зоне синапсов, расположенных на дендритном стволе. Эта ситуация наблюдается, в основном, в ранние периоды развития нейронов, когда большинство синапсов располагаются непосредственно на дендритном стволе и формируются раньше, чем вырастают дендритные филлоподии. В дальнейшем количество таких «ранних» синапсов падает, а число зрелых аксо-шипиковых синапсов увеличивается, то есть присутствие в районе формирования шипиков элементов формирующегося синапса является важным индуцирующим сингналом для формирования структуры шипиков и зрелого аксо-шипикового синапса.
3. Формирование шипиков без индукционного воздействия формирующихся синапсов. Шипики на дендритах могут формироваться и раньше, чем в этом районе сформируются синаптические контакты. Так происходит, например, при формировании шипиков на дистальных веточках дендритной системы клеток Пуркинье мозжечка.
Аксон. Аксон – обычно самый длинный отросток нейрона, нередко интенсивно ветвящийся в своих терминальных отделах. Одной из отличительных черт аксона является наличие т.н. «возвратных коллатералей» - терминальных веточек аксона, образующих аксо-аксональные контакты на собственном нейроне. Они имеют важное регулирующее влияние (в основном тормозное) на работу аксона. Из-за большой длины аксона его аксоплазма может значительно превосходить общий объем цитоплазмы перикариона и дендритов нейрона вместе взятых. Принципиальное отличие аксона от остальных отделов нейрона заключается и в отсутствии в его
цитоплазме мРНК и рибосом и наличии (в спайк-генерирующих нейронах) в плазматической мембране большого числа Na-ионных каналов, участвующих в генерации и проведении потенциала действия.
В последнее время показано, что в растущих веточках аксона (конусах роста) и при регенерации в аксоне осуществляется локальный синтез белков. Компоненты белоксинтетического аппарата (субъединицы рибосом и сами рибосомы, мРНК, необходимые ферменты, аминокислоты) поставляются в районы синтеза аксональным транспортом. Неодинаковая структура цитоплазмы аксона и дендритов связаны, как полагают, и с отличиями в расположении элементов цитоскелета (см. ниже).
Место отхождения аксона от тела нейрона называется аксонным холмиком, который, суживаясь, постепенно переходит в начальный сегмент аксона. Именно эти начальные участки аксона являются местом генерации потенциала действия и отличаются большим числом ионных каналов в плазмолемме. Число рибосом в аксонном холмике невелико, и по мере сужения отростка они полностью исчезают. В начальном сегменте аксона рибосомы находятся только около постсинаптических активных зон аксо-аксональных синапсов. Этот сегмент характеризуется тем, что микротрубочки цитоскелета здесь собраны в небольшие группы и около плазмолеммы имеется слой электронно-плотного материала. Слой субплазмолеммального материала находится также в перехватах Ранвье. Это связано, по-видимому, со множеством ионных каналов, особенно натриевых, в плазматической мембране. В аксоне в большом количестве находятся элементы агранулярной эндоплазматической сети (АЭС) – важной транспортной системы отростков.
Морфология дендритов.
Нейроны имеют две системы отростков: аксон и дендриты и одной из задач нейроморфологии является определении ключевых различий или сходства между ними.
На светооптическом уровне исследования при изучении срезов нервной ткани, окрашенной солями серебра по методу Гольджи, видны некоторые морфологические отличия аксона от дендритов. Они в ряде случаев не абсолютны, но теме не менее позволяют проанализировать имеющийся нейрогистологический материал. (табл ).
Табл. Ххх Некоторые сравнительные характеристики аксона и дендритов нервных клеток позвоночных
Свойства или признаки |
Аксон |
Дендриты |
Примечания |
Количество на нейрон |
Всегда один (или отсутствует) |
Как правило несколько (может быть один дендрит и отсутствовать) |
|
Длина отростков |
От нескольких микрон до метра и более |
От нескольких микрон до нескольких сот микрон |
|
Характер отхождения отростка от нейрона |
Преимущественно от тела нейрона, иногда от крупных дендритов |
От тела нейрона |
|
Степень разветвленности |
Самая разнообразная |
Самая разнообразная |
Разветвленность аксона, как правило, превышает таковую у дендритов данной клетки |
Характер ветвления боковых веточек (коллатералей) |
Отходят преимущественно под прямым углом, встречаются возвратные ветвления и сильно извитые |
Отходят преимущественно под острым углом по ходу основной ветви, возвратные коллатерали редки. |
|
Стабильность ветвлений |
Более вариабелен среди нейронов данного типа |
Более стабилен |
|
Ниличие шипиков |
Есть, много и разного вида* |
Нет |
* Некоторые типы клеток не имеют на дендритах шипиков (бесшипиковые нейроны) |
Наличие рибосом и ШЭР |
Рибосомы есть по всей длине дендрита, ШЭР есть в проксимальных сегментах дендритов и в узлах ветвления |
Нет * |
* Получены данные о наличии в аксонах и синапсах рибосом и механизма синтеза белка |
Наличие цистерн ГЭР |
Есть по всей длине аксона |
Есть по всей длине дендритов |
|
Наличие миелиновой и немиелиновой оболочек |
Есть оба вида оболочек |
Есть миелиновая оболочка (но редко и очень тонкая) |
|
Диаметр отростков |
Уменьшается по мере удаления от тела клетки и ветвления |
Практически постоянный по всей длине аксона |
|
Преобладающая ориентация и количество микротрубочек |
Однонаправленная (от клетки к синаптическому окончанию), в крупных аксонах меньше, чем в дендритах |
Разнонаправленная, особенно много в крупных дендритах и их ветвях |
|
Скорость проведения импульса и направление* |
До 120 м/сек, в обоих направлениях |
До 1 м/сек, в обоих направлениях |
* В каждый данный момент времени по аксону или дендритам импульс идет в одном направлении |
Способность к генерации, проведению и передачи нервного импульса |
есть |
Есть, но проведение возбуждения по дендриту идет в уменьшительной форме (угасания)* |
* Дендриты способны генерировать потенциал действия |
|
|
|
|
|
|
|
|
Аксон и дендриты имеют помимо перечисленных выше морфологических и функциональных различий набор молекулярно-биохимических маркеров, присутствие которых на мембранах или в цитоплазме отростков во многом определяет их специфику и свойства.
Цитоскелет нейрона. Аксонный и дендритный транспорт
На светооптическом уровне в нейроне и его отростках при окраске солями серебра выявляется тонкая сеть нитей, толщиной 0,5 – 3 мкм, и названных «нейрофибриллами». Оказалось, что это пучки цитоскелетных фибрилл разных видов, склеивающихся в пучки под действием фиксации.
Цитоскелет нервной клетки имеет большое значение в жизнедеятельности нейронов, и, как и в других клетках животных и человека, состоит из микротрубочек, промежуточных филаментов и микрофиламентов.
Микротрубочки.
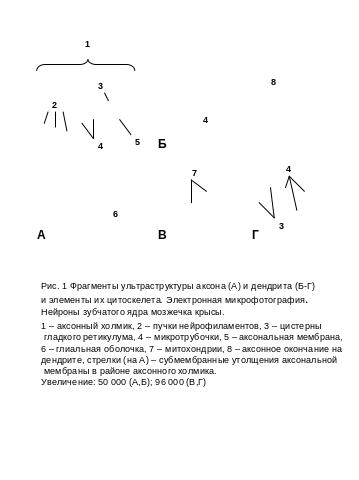
Большинство микротрубочки формируются из белка тубулина в цитоплазме в т.н. «микротрубочкоорганизующем центре, МТОЦ», расположенном в районе клеточного центра (центриоли). Стенка микротрубочек состоят из 13 концентрически расположенных глобул белка тубулина. Каждая молекула тубулина является димером и состоит из α и ß-тубулина. Диаметр микротрубочки постоянен и составляет 24 нм по наружному краю и 15 нм по внутреннему контуру. Длина микротрубочек может быть самой разной, от нескольких десятков нанометров до десятков микрон. Это зависит от типа нервной клетки, места расположения микротрубочек в нейроне и отростках. В нейроне микротрубочки присутствуют в двух формах – длинные стабильные, и, как правило, неподвижные микротрубочки и короткие подвижные микротрубочки. В нейронах с помощью специальных ферментов – катанина и спастина – происходит трансформация микротрубочек из одного вида в другой. Катанин разрезает длинные стабильные микротрубочки на короткие мобильные фрагменты (примерно 10нм в длину), которые затем могут перемещаться по цитоплазме и отросткам нейрона на десятки и сотни микрон, после чего короткие фрагменты микротрубочек, возможно при участии спастина, опять могут собираться в длинные стабильные формы. (Рис. 1).
Каждая микротрубочка имеет быстрорастущий «+» – конец, где идет активная сборка новых фрагментов и «-» – конец, где рост микротрубочки блокируется специальными «кэпирующими» белками, что способствует росту микротрубочки на (+) конце. В теле нейрона основная масса микротрубочек ориентирована радиально по направлению от МТОЦ (минус конец) к периферии клетки (плюс конец). Часть микротрубочек в цитоплазме нейрона может быть ориентирована и в противоположном направлении. В отростках нейрона микротрубочки располагаются, как правило, упорядоченно и вдоль длинной оси отростков. Среднее расстояние между соседними отдельными микротрубочками составляет от 20 до 60 нм. (Рис. 2).
В аксоне встречается несколько разновидностей микротрубочек. Большинство располагается поодиночке вдоль длинной оси аксона и их плюс конец направлен в сторону аксонного окончания (синапса). В месте отхождения аксона от тела клетки, в

т.н. «аксоном холмике», микротрубочки образуют компактные пучки из 10-25 штук, также ориентированные к периферии аксона. ( Рис. 2, а). Именно здесь происходит сортировка материала, транспортируемого дальше по аксону. В аксоне микротрубочки более стабильны и менее подвержены воздействию различных факторов, чем в теле нейрона и в дендритах. В районе синапса обнаружены особая разновидность микротрубочек - «изогнутые микротрубочки» – они принимают участие в транспорте синаптических пузырьков с медиаторами непосредственно к пресинаптической мембране.
В дендритах микротрубочки (Рис. 2 б-г) располагаются вдоль оси отростка, но ориентация их концов может быть противоположная по отношению друг к другу. Это характерно, правда, только для проксимальных участков дендритов (в дистальных участках «+» - конец микротрубочек направлен к периферии).
Важным элементом структуры микротрубочек, определяющим во многом их свойства, является наличие большого количества специализированных микротрубочко-ассоциированных протеинов (МАР-белки). Выделяют два основных типа этих белков: 1) высокомолекулярные МАР-белки нескольких классов (МАР1-5); 2) низкомолекулярные тау-протеины (некоторые виды последних обнаруживаются только в нейронах). Роль МАР белков в организации цитоскелета нервной ткани очень важна: они обеспечивают стабильность микротрубочек, контролируя процессы сборки и разборки, связывают микротрубочки друг с другом и с другими компонентами цитоскелета, а также с плазматической мембраной и органоидами клетки. Именно различия в структуре МАР – белков определяют специфику микротрубочек в теле нейрона, аксоне и дендритах, поскольку строение самих микротрубочек везде одинаковое. В качестве примера можно привести белок МАР-2a,b, который присутствует только в дендритах, тогда как белок МАР – 3 есть только в аксонах и глии. Если заблокировать синтез тау-белков в культуре нейрональных клеток, то они теряют свои аксоны, сохраняя только дендриты. Введение генов тау-протеинов в мутантные нервные клетки, которые не экспрессируют этот белок, приводит к активному росту клеточных отростков.
Все процессы, связанные с образованием микротрубочек, их подвижностью и участием в клеточных процессах идут с затратой энергии молекул ГТФ и ГДФ. Стабильность микротрубочек связана с рядом как внутренних, так и внешних факторов. Среди внешних следует отметить такие как: уровень ионов Са+2 и Мg+2 в нейроне, температуру (чем ниже температура, тем ниже темп сборки микротрубочек и скорость транспорта), уровень содержания кислорода в мозгу, РН среды (чем выше РН, тем интенсивнее идут процессы распада микротрубочек) и другие. Средний срок полужизни микротрубочки в нейроне ~ 10 - 20 минут.
К блокаде полимеризации или деполимеризации микротрубочек и, как следствие, нарушению транспортных процессов в нейронах приводит воздействие таких веществ - цитостатиков, как: колхицин, колцемид, винпрестин, винбластин, нокадазол, таксол. Их используют при химиотерапии опухолей, чтобы заблокировать деление раковых клеток.
Таким образом, в нейроне и его отростках микротрубочки пребывают в постоянном процессе сборки, разборки и перемещении по цитоплазме нейрона. Такое состояние микротрубочкового скелета клетки получило наименование – «динамическая нестабильность цитоскелета».
Нейрофиламенты (промежуточные филаменты).
У человека более 65 генов связаны с синтезом филаментозных белков. Соединяясь друг с другом, отдельные нейрофиламентозные белки (мономеры) формируют в нервных клетках вначале гомодимеры из двух фибрилл, которые затем соединяются попарно и образуют зрелую протофибриллу – гомотетрамер, который состоит из четырех идентичных молекул белка. Далее происходит полимеризация нейрофибриллярных протофибрилл в зрелую нейрофибриллу, диаметром ~ 10 нм, и состоящую из 8 длинных протофибрилл. Нейрофиламенты представлены тремя нейроспецифическими белками: NF-L, NF-Н, NF- M. и являются своеобразной «визитной карточкой» нейронов, т.к. они есть только в нервных клетках или клетках общего с ними происхождения.
Сборка нейрофиламентов происходит достаточно быстро. В экспериментах in vitro показано, что в течение первых секунд формируются нейрофиламенты длиной в 60нм, за первую минуту – 300нм, а через 15-20 минут длина увеличивается до 0,5 – 1 микрона. Процесс удлинения на этом не оканчивается, и после нескольких часов сборки мы имеем очень длинные нейрофиламенты. Ориентированы нейрофиламенты преимущественно вдоль длинной оси отростков нейрона. Они могут быть как одиночными, так и образовывать пучки. Особенно их много в районе аксонного холмика. В нейронах ЦНС при болезни Альцгеймера, рассеяном склерозе и другой патологии отмечается резкое увеличение концентрации нейрофиламентов и нарушение их ориентации при отчетливом уменьшении концентрации микротрубочек (Рис.3).
Нейрофиламенты – структуры более стабильные, чем микротрубочки (среднее время полужизни нейрофиламентов составляет ~ 40 мин.). Тем не менее, они также находятся в состоянии «динамической нестабильности», постоянно разбираясь и собираясь в теле и отростках нейрона, с помощью специальных ферментов. В районе синаптического окончания нейрофиламентов нет – в пресинаптической области они разрушаются и их компоненты с помощью обратного транспорта возвращаются в аксон и тело нейрона.
В целом промежуточные филаменты выполняют в нейроне механическую функцию, поддерживая форму тела и отростков. Они участвуют в росте и регенерации отростков, а также являются важным компонентом внутриклеточного транспорта. Нейрофиламенты тесно связаны друг с другом, с микротрубочками, клеточной и аксональной мембраной и другими компонентами клетки, образуя сложную трехмерную цитоскелеткную сеть в теле и отростках нейронов.
Микрофиламенты (актиновые фибриллы).
Микрофиламенты состоят из высококонсервативного внутриклеточного белка – актина. Они имеют диаметр 5-8 нм и образованы из двух нитей фибриллярного актина (F-актин), который образуется путем полимеризации молекул глобулярного актина (G-актина). У человека обнаружено несколько изоформ G-актина, кодируемые более чем 30-ю генами и объединенными в три суперсемейства: α - актины, β – актины и c –актины. В нервной ткани встречаются преимущественно β – и c –актины. Регулируемая полимеризация G-актива происходит в клетке в присутствии катионов Mg+2, Ca+2 и K+. Актиновые фибриллы располагаются повсеместно в теле и отростках нейрона, образуя разветвленную сеть. Особенно много актиновых фибрилл в аксоном холмике и ножках шипиков.
Рост фибриллы идет с двух концов, но на «+» - конце полимеризация идет сильнее (за секунду примерно 70 молекул актина присоединяется и 2 молекулы отсоединяется), тогда как на «-» конце, медленнее (20 молекул присоединяется, а одна отсоединяется).
В состав актиновой фибриллы входит целый ряд вспомогательных «актинсвязывающих белков». Они позволяют образовывать в цитоплазме из отдельных актиновых фибрилл целую сеть актиновых скелетных нитей, а также связывают актин с плазматической мембраной или с другими компонентами цитоскелета. Взаимодействуя с миозиновыми молекулами в цитоплазме нейрона и отростков, актиновые филаменты формируют механо-химический комплекс (подобный таковому в мышечных волокнах), участвуя в образовании и росте отростков, ретрактильной подвижности клеток, и обеспечивая транспортные процессы в нейроне.
Аксонный и дендритный транспорт.
В нейроне, как и в других клетках организма, постоянно происходят процессы распада молекул, органоидов, других компонентов клетки. Их необходимо постоянно обновлять. Нейроплазматический транспорт важен для обеспечения электрических и неэлектрических функций нейрона, для осуществления обратной связи между отростками и телом нейрона. При повреждении нервов необходима регенерация поврежденных участков и восстановление иннервации органов.
Разнообразные вещества транспортируются по отросткам нейрона с разной скоростью, в разных направлениях и с использованием разных механизмов транспорта. Выделяют два основных вида транспорта: прямой (антероградный) – от тела клетки по отросткам к их периферии и обратный (ретроградный) – по отросткам нейрона к телу клетки.
(табл. 1)
Табл. 1 Основные компоненты аксонного и дендритного транспорта в нейронах позвоночных (по данным разных авторов)
Компоненты и субкомпоненты транспорта |
Скорость мм/сутки |
Что транспортируется |
Морфологический субстрат транспорта |
Прямой (антероградный) аксональный транспорт |
|||
Быстрый (Fast) |
|||
I |
200- 500
|
Медиаторы и их предшественники, ферменты синтеза медиаторов, белки плазматической мембраны, мембранные органоиды, нейрогормоны, |
Синаптические пузырьки, цистерны гладкого ретикулума, нейросекреторные гранулы, цитоскелетная сеть |
Промежуточный |
|||
II |
50 - 100 |
Белки митохондрий, липиды мембран |
Митохондрии, цитоскелет |
III |
15 |
Миозиновые белки, |
Цитоскелет |
Медленный (Slow) |
|||
IV SCb |
2- 4 |
Актин, клатрин, актинсвязывающие белки, ферменты метаболизма нейрона, белки аксоплазмы |
Цитоскелет (микротрубочки, микро- и нейрофиламенты), микротрабекулярная сеть |
V SCa |
0,2- 1 |
Белки нейрофиламентов, тубулин и фрагменты микротрубочек, ферменты аксоплазмы |
Цитоскелет (микротрубочки, микро- и нейрофиламенты), микротрабекулярная сеть |
Прямой быстрый дендритный транспорт |
|||
I D |
200- 400 |
Белки постсинапса, рецепторные комплексы, белки цитоплазмы и мембран дендрита и шипиков |
Цитоскелет, гладкий ретикулум, транспортные пузырьки |
Обратный (ретроградный) транспорт |
|||
I R |
100- 300 |
Отработанные лизосомы и митохондрии, ростовые и трофические факторы, вирусы. |
Мультивезикулярные и мультиламеллярные тела, цитоскелет, эндосомы |
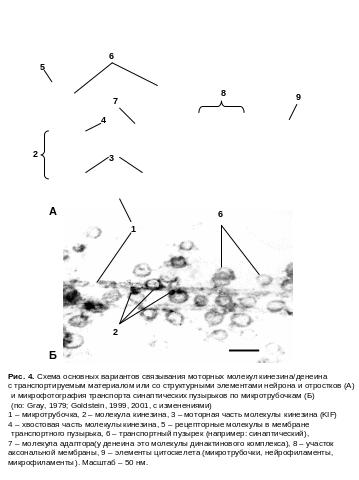
В осуществлении транспортных процессов в нейроне участвуют пять групп «моторных» белков, тесно связанных с цитоскелетной сетью. В их состав входят такие белки как кинезины, денеины и миозины.
В осуществлении транспортных процессов в нейроне участвуют пять групп т.н. «моторных» молекул (Рис. xx ).
1-3 Группа. Кинезины
В составе этой группы выделяют три типа кинезиновых белков.
1. Группа. Конвекционный кинезин (kinesin - I или KIF-5). Он был идентифицирован в нервной системе головоногих моллюсков и млекопитающих в 1985 году, а затем и в клетках других животных, включая низших эукариот. Он тесно связан с микротрубочками и является одним из самых главных транспортных белков клетки, осуществляя транспорт материалов (cargo) вдоль микротрубочек по направлению к ее плюс концу. С его помощью транспортируются в отростках нейронов митохондрии, лизосомы, цистерны эндоплазматического ретикулума, синаптические пузырьки, а также ряд немембранных комполнентов клетки (молекулы и-РНК, белки и фибриллы нейрофиламентов).
Состоит молекула кинезина -1 из двух тяжелых и двух легких полипептидных цепей. Из тяжелых и легких цепей каждая кодируются тремя генами. Легкие и тяжелые цепи могут комбинироваться в различных сочетаниях и, как полагают, могут, таким образом, формировать различные разновидности молекул кинезина – I, транспортирующие при этом разные компоненты внутри клетки.