

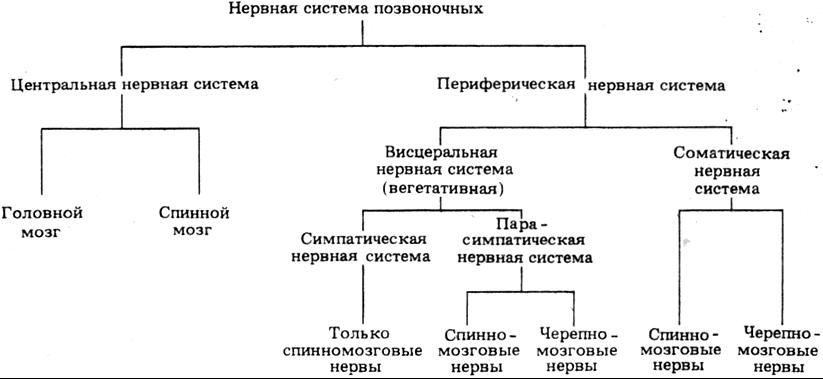
Нервная система
Нервная система 1
Этапы развития нервной системы 3
Биоэлектрическия явления на мембранах клеток как общий принцип работы НС. 4
Потенциал покоя (ПП) 4
(к оглавлению) 4
Возбудимость и раздражимость 5
Локальное возбуждение (рецепторный потенциал – РП) захватывает лишь отдельные структуры возбудимых образований. Оно характеризуется следующими особенностями: 6
Потенциал действия 6
Ключевые понятия: 6
Механизм развития. В клетках нервной и мышечной ткани, имеющих потенциал-зависимые каналы, возбуждение развивается следующим образом (мВ – милливольты, мс – миллисекунды; отсчет времени начинается с момента воздействия раздражителя): 6
Механизм работы φ-зависимых каналов. 7
Поэтапный анализ развития ПД и изменения возбудимости 8
(к оглавлению) 9
Нервный импульс. 10
Генерация и передача возбуждения в рецепторах 10
Распространение нервного импульса 10
Синаптическая передача возбуждения 11
Разновидности синапсов: 11
Химические синапсы 11
а) Нервно-мышечное (мионеврального) соединение. 11
б) Химические межнейронные синапсы. 12
Торможение, возникающее в нервно-мышечных или нервно-железистых соединениях, называется периферическим, а реализуемое в структурах ЦНС — центральным. Явление центрального торможения было открыто в 1862 г. И. М. Сеченовым. Дальнейшую разработку теории торможения существенный вклад внесли Н. Е. Введенский, Ч. Шеррингтон, А. А. Ухтомский и др. 12
Интеграция нейронных связей 13
Принципы построения нейронных сетей. 13
14
(к оглавлению) 14
Центральная нервная система 15
Спинной мозг 15
Рефлекторная функция спинного мозга 15
Проводниковые функции спинного мозга 16
А. Восходящие пути 17
Б. Нисходящие пути 17
Столбы сп. мозга 17
Физиологическое значение 17
Головной мозг 17
Ретикулярная формация. 18
Структурные отделы головного мозга 18
I. Продолговатый мозг 18
II. Задний мозг 18
Мозжечок 18
Мост 18
III. Средний мозг 19
IV. Промежуточный мозг 19
V. Конечный мозг 19
Базальные ганглии. 19
Большой мозг. 19
Функциональные зоны коры. 20
Лимбическая система. 21
Методы исследования функций головного мозга 22
Периферическая нервная система 23
Рефлексы и рефлекторные дуги 23
Вегетативная нервная система 24
Различия между симпатической и парасимпатической системами 25
Эффекты симпатической и парасимпатической систем в организме 25
Черепномозговые нервы млекопитающих, их функции и иннервируемые органы 25
Этапы развития нервной системы
Появление многоклеточных организмов явилось первичным стимулом для дифференциации систем связи, которые обеспечивают целостность реакций организма, взаимодействие между его тканями и органами. Это взаимодействие может осуществляться гуморальным путем посредством поступления гормонов и продуктов метаболизма в тканевую жидкость, кровь и лимфу и за счет функции нервной системы, которая обеспечивает быструю передачу возбуждения, адресованного к вполне определенным мишеням. Видимо, обе системы (эндокринная и нервная) развивались параллельно.
Нервная система беспозвоночных животных
Нервная система на пути структурного и функционального развития проходит через несколько этапов, которые у первично- и вторичноротых животных серьезно различаются.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у кишечнополостных. Их нервная сеть представляет собой скопление мультиполярных и биполярных нейронов, отростки которых могут перекрещиваться, прилегать друг к другу, и лишены функциональной дифференциации на аксоны и дендриты. Диффузная нервная сеть не разделена на центральный и периферический отделы.
Дальнейшее развитие нервной системы идет по пути централизации и концентрации нервных элементов. Лестничный тип нервной системы плоских и круглых червей – одна или несколько пар продольных нервных стволов, соединенных большим числом поперечных и кольцевых комиссур. Концентрация нервных элементов сопровождается их погружением вглубь тела.
Плоские черви являются билатерально симметричными животными с четко выраженной продольной осью тела. Движение свободноживущих форм осуществляется преимущественно в сторону головного конца, где концентрируются рецепторы, сигнализирующие о приближении источника раздражения. К числу таких рецепторов турбеллярий относятся пигментные глазки, обонятельные ямки, статоцист, чувствительные клетки покровов, наличие которых способствует концентрации нервной ткани на переднем конце тела. Это приводит к формированию головного ганглия (процесс энцефализации).
Ганглионизация нервных элементов получает дальнейшее развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У примитивных кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев соединены между собой поперечными комиссурами. С другой парой, расположенной в соседнем сегменте они соединены коннективами, т.е. сохраняется лестничный тип нервной системы, но в усложненном варианте.
У более продвинутых кольчатых червей наблюдается сближение брюшных стволов вплоть до полного слияния ганглиев правой и левой сторон и перехода от лестничной к цепочечной нервной системе (брюшная нервная цепочка с окологлоточным кольцом). Идентичный, цепочечный тип строения нервной системы существует и у низших членистоногих с различной выраженностью концентрации нервных элементов, которая может осуществляться не только за счет слияния соседних ганглиев одного сегмента, но и при слиянии последовательных ганглиев различных сегментов.
В дальнейшем, процесс ганглионизации у беспозвоночных приводит к формированию нервной системы узлового типа (ганглионарный тип), которая характерна высшим насекомым и высшим моллюскам. У продвинутых головоногих, слившиеся ганглии формируют дифференцированный на отделы «мозг».
Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением. Низший уровень интеграции в сегментарных ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время «мозг» представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значимости информации.
Нервная система позвоночных животных
Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая в процессе онто- и филогенеза дифференцируется на различные отделы. У наиболее древних хордовых (бесчерепных) головной мозг отсутствует, и нервная трубка представлена в малодифференцированном состоянии.
В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной — процесс энцефализации, который был рассмотрен выше на примере беспозвоночных животных. В период филогенетического развития от уровня бесчерепных до уровня круглоротых формируется головной мозг. Исследование ЦНС современных круглоротых показывает, что их головной мозг в зачаточном состоянии содержит все основные структурные элементы. Развитие вестибулярной системы, возникновение дыхательного центра создают основу для формирования заднего мозга. Развитие зрительной рецепции дает толчок к закладке среднего мозга. На дорсальной поверхности нервной трубки развивается зрительный рефлекторный центр — крыша среднего мозга. И наконец, развитие обоняния способствует формированию переднего, или конечного, мозга, к которому примыкает слаборазвитый промежуточный мозг.
Последующее развитие мозга идет в различных классах позвоночных и даже внутри одного и того же класса по-разному, но во всех случаях исходная трехчастная структура утрачивается, так как передний, средний и задний мозг подвергаются дальнейшему подразделению. Мозг взрослого позвоночного состоит из пяти отделов, значение которых у представителей каждого класса зависит от образа жизни и уровня структурной и функциональной сложности, достигнутой этим классом. Некоторые участки мозга увеличиваются в размерах, что указывает на их особую важность, тогда как другие уменьшаются.
Р
 ыбы
в поисках пищи сильно зависят от
обоняния, и у них большие обонятельные
доли, перерабатывающие сенсорные
сигналы. Крупный мозжечок осуществляет
у них координацию движений, а весьма
развитые зрительные доли мозга хотя и
участвуют в зрительных реакциях, но в
основном служат главным координирующим
центром мозга. Зрительные доли составляют
главную часть мозга и у амфибий, у которых
они выполняют ту же роль, что и у рыб. У
рептилий размеры среднего мозга
уменьшаются, а переднего - увеличиваются.
Эта тенденция продолжается и у птиц, у
которых увеличиваются размеры
таламуса и полосатых тел, координирующих
сложные инстинктивные формы поведения.
И наконец, для мозга млекопитающих
характерно наличие больших полушарий
и хорошо развитый таламус. Эти две
структуры, содержащие ядра и проводящие
пути, отражают потребность млекопитающих
в хранении сенсорной информации и в
интегрировании всех произвольных
действий.
ыбы
в поисках пищи сильно зависят от
обоняния, и у них большие обонятельные
доли, перерабатывающие сенсорные
сигналы. Крупный мозжечок осуществляет
у них координацию движений, а весьма
развитые зрительные доли мозга хотя и
участвуют в зрительных реакциях, но в
основном служат главным координирующим
центром мозга. Зрительные доли составляют
главную часть мозга и у амфибий, у которых
они выполняют ту же роль, что и у рыб. У
рептилий размеры среднего мозга
уменьшаются, а переднего - увеличиваются.
Эта тенденция продолжается и у птиц, у
которых увеличиваются размеры
таламуса и полосатых тел, координирующих
сложные инстинктивные формы поведения.
И наконец, для мозга млекопитающих
характерно наличие больших полушарий
и хорошо развитый таламус. Эти две
структуры, содержащие ядра и проводящие
пути, отражают потребность млекопитающих
в хранении сенсорной информации и в
интегрировании всех произвольных
действий.
Относительные размеры продолговатого мозга в ходе эволюции позвоночных менялись мало, и это говорит о существенной роли, которую играет продолговатый мозг в рефлекторной регуляции всех жизненно важных функций, таких, например, как работа сердца. (к оглавлению)
Биоэлектрическия явления на мембранах клеток как общий принцип работы НС.
В 18 веке, проводя опыты на нервно–мышечных препаратах лягушки, Гальвани обнаружил, что если к препарату приложить две соединенные между собой пластинки из разнородных металлов (при этом одна пластинка касается мышцы, а другая – нерва), то мышца сокращается. Гальвани со своим племянником, физиком Джованни Альдини, объяснил это явление протеканием «животного электричества», которое, по их мнению, зарождалось в нервах и запасалось в мышцах. Они предположили, что от мышцы к нерву через металлический проводник перетекали некие «электрические флюиды» и что именно этот разряд, идущий из мышцы, вызывал ее сокращение. Эти положения были высказаны в 1791 году, и с высоты сегодняшних знаний они, конечно, во многом представляются неверными. Однако работа Гальвани и Альдини, появившаяся в этот революционный век, побудила многих исследователей посвятить свои труды двум важным областям человеческого знания: физиологии возбуждения нервов и мышц и химической природе электричества. В XIX веке стало очевидно, что сигналы, возникающие в нервных клетках и других возбудимых тканях, обусловлены электрическими свойствами клеточных мембран, а конкретнее – перераспределением ионов по две стороны от мембран.
Для изучения электрических процессов в живой ткани можно ввести в эту ткань два электрода и измерять потенциал, создаваемый в результате протекания тока через внеклеточную жидкость. Но, поскольку, разность зарядов, порождающая эти токи, возникает, прежде всего, по разные стороны клеточной мембраны, более прямую и точную количественную оценку биоэлектрических явлений можно получить, измеряя трансмембранные токи в отдельных клетках. Для этого необходимо сравнить электрический потенциал цитоплазмы, с потенциалом жидкостной внешней среды. Разность этих потенциалов называется мембранным потенциалом. Для измерения мембранного потенциала один регистрирующий стеклянный микроэлектрод помещают во внеклеточную жидкость, а другой – во внутриклеточную среду. Разность потенциалов между этими электродами усиливают с помощью электронного усилителя и выводят на регистрирующее устройство. Стеклянный микроэлектрод очень мал, и представляет собой трубочку, заполненную раствором электролита (например, KCl). Мембранным потенциалом называют разность между внутриклеточным потенциалом (регистрируемым микроэлектродом) и внеклеточным (регистрируемым серебряной проволочкой, помещенной в окружающую клетку среду). Внеклеточный потенциал условно считают равным нулю. Фактически же электронный усилитель вычитает внеклеточный потенциал из внутриклеточного и регистрирует их разность.
Потенциал покоя (ПП)
Каждая клетка в невозбужденном состоянии (в состоянии покоя) характеризуется наличием трансмембранной разности потенциалов. Обычно этот так называемый потенциал покоя составляет от – 30 до –100 мВ (отрицательный заряд на внутренней поверхности мембраны), хотя у некоторых клеток он может быть и ниже. Его величина зависит от типа клетки и ионного состава окружающей среды. В создании потенциала покоя участвуют два фактора. Первый из них – это неравномерное распределение неорганических ионов между клеточным содержимым и наружной средой. Такое распределение обусловлено активным транспортом ионов с помощью белковых мембранных насосов (в данном случае – «Na+–K+ - насос»). Второй фактор – это наличие в клеточной мембране белковых ионных каналов, пропускающих лишь некоторые из присутствующих в клетке и окружающей среде ионов. В данном случае это Na+– каналы (в покое закрыты) и K+– каналы (в покое открыты). Мы уже знаем, что неравномерное распределение ионов порождает их диффузионный поток по градиенту концентрации (ΔС). Именно эти механизмы ответственны за наличие потенциала покоя.
Создание потенциала покоя:
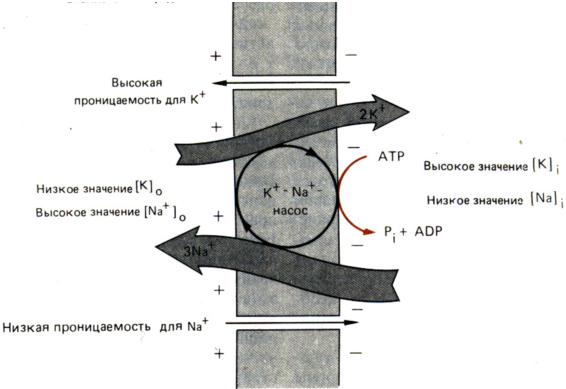 Na
+–K
+
– насос работает таким образом, что за
один цикл, удаляя из клетки три иона
Na+,
он вводит в нее два иона К+,
т.е. в конечном счете удаляет из клетки
положительные заряды (прямой вклад в
создание потенциала покоя). Но создаваемый
с помощью натриевого насоса потенциал
редко превышает несколько милливольт,
т.е. крайне незначителен.
Косвенная роль Na+–К+–
насоса связана с тем, что он поддерживает
высокую
концентрацию
К+
и низкую концентрацию Na+
во
внутриклеточной среде.
Na
+–K
+
– насос работает таким образом, что за
один цикл, удаляя из клетки три иона
Na+,
он вводит в нее два иона К+,
т.е. в конечном счете удаляет из клетки
положительные заряды (прямой вклад в
создание потенциала покоя). Но создаваемый
с помощью натриевого насоса потенциал
редко превышает несколько милливольт,
т.е. крайне незначителен.
Косвенная роль Na+–К+–
насоса связана с тем, что он поддерживает
высокую
концентрацию
К+
и низкую концентрацию Na+
во
внутриклеточной среде.
Основным же фактором, ответственным за создание потенциала покоя, служит высокая проницаемость мембраны для калия и низкая для натрия.
а) Как уже было сказано, в покое Na+– каналы закрыты, и ионы натрия не могут войти в клетку (по ΔС), в результате его концентрация в клетке значительно ниже, чем в окружающей среде (более чем в 10 раз).
б) K+– каналы открыты, и калий диффундирует из клетки (по ΔС) до тех пор, пока его выходу не будет препятствовать значительный отрицательный заряд цитоплазмы.
Итогом работы Na+–K+ – насоса и состояния ионных каналов является отрицательный заряд внутренней поверхности плазмалеммы – потенциал покоя (ПП). Важно понять, что активный транспорт ионов через мембрану, осуществляемый Na+–K+ – насосом, не отвечает непосредственно за создание потенциала, но необходим для поддержания ионных концентрационных градиентов, которые ответственны за возникновение мощных диффузионных потоков.
(к оглавлению)
Возбудимость и раздражимость
Раздражимость – один из атрибутов жизни и, следовательно, характерная черта всех биологических систем. Она выражается в способности живых клеток, тканей или целого организма реагировать на внешние или внутренние воздействия (раздражители) и лежит в основе приспособления к изменяющимся условиям среды. Раздражимость сопровождается комплексом неспецифических изменений, выражающихся в сдвигах обмена веществ, состояния протоплазмы, а у высокоорганизованных животных связана с выполнением определенных функций.
У животных, не имеющих нервной системы, реакции на раздражения охватывают всю протоплазму и выражаются главным образом в форме двигательных реакций (таксисов).
Раздражимость у растений обусловлена структурными и функциональными изменениями мембран, тургорного давления и лежит в основе их регуляторной системы. Наиболее ярко она проявляется в реакциях на свет (фототропизм, фотопериодизм), на гравитационное поле (геотропизм), в двигательных реакциях (настии).
Способность нервных и мышечных клеток отвечать на раздражение называется возбудимостью.
Возбудимость (в физиологии) как одна из форм раздражимости возникла в процессе эволюции в связи с развитием специфических возбудимых тканей и является исключительно их свойством. Возбудимыми тканями являются нервная, мышечная и, в некоторой степени, железистый эпителий. Возбудимость – это способность воспринимать воздействия раздражителей и отвечать на них реакцией возбуждения. Возбуждение связано со специфической чувствительностью клеточных мембран этих тканей, с их способностью отвечать на действие адекватных раздражителей (см. ниже) соответствующими изменениями ионной проницаемости и мембранного потенциала (активный электрический ответ). Например: вернемся к схеме формирования ПП и представим, что, от сигнала раздражителя, закрытые в покое Na+– каналы открылись. Na+ начнет перемещаться в клетку под воздействием двух сил: а) градиента концентраций (по ΔС), б) электростатического притяжения. Это приведет к смене отрицательного заряда внутренней поверхности мембраны на менее отрицательный и даже положительный, т.е. к деполяризации мембраны.
Различают общие и специфические признаки возбуждения. К общим для всех возбудимых тканей признакам относят изменение мембранного потенциала и сопутствующие ему изменения уровня обменных процессов с выделением энергии (тепловой, электрической, лучистой). При этом раздражитель (стимул) служит лишь пусковым фактором: его энергия может быть значительно ниже, чем получаемая за счет обмена веществ энергия возбуждения.
Специфические признаки возбуждения определяются переходом от состояния покоя к свойственной деятельности. Нервная ткань генерирует и проводит импульсы, мышечная сокращается, железистая образует и выделяет секреты.
Интенсивность, длительность и быстрота реакций в ответ на раздражения неодинаковы для различных возбудимых тканей.
Раздражители по своей энергетической сущности могут быть механическими, химическими, термическими, электрическими и т.д., а по биологическому значению — адекватными и неадекватными.
Адекватные (соответствующие, специфические) — это раздражители, способные при минимальной энергии раздражения вызывать возбуждение клеток, специально приспособленных для восприятия данного вида раздражителя. Так, для клеток сетчатки глаза адекватный раздражитель — кванты света, для слуховых рецепторов — звуковые колебания и т.п.
Неадекватные (общие, неспецифические) раздражители также вызывают ответную реакцию возбудимых структур, но лишь при значительной силе и длительности воздействия (например, ощущение от вспышки света при сдавливании глазного яблока). Электрический ток считают адекватным раздражителем для возбудимых тканей, поскольку их функциональная активность всегда сопровождается биоэлектрическими явлениями.
Различие свойств возбудимых и невозбудимых тканей связано с тем, что клеточные мембраны могут реагировать на электрические раздражители двумя совершенно разными способами: пассивным и активным.
1. Пассивный электрический ответ – это изменение мембранного потенциала, связанное с тем, что через мембрану начинает течь электрический ток от какого–либо постороннего источника.
Подобное изменение потенциала не зависит от молекулярных процессов в самой мембране (например, от срабатывания ионных каналов). Этим пассивный ответ отличается от активного (возбуждения). Пассивный ответ возникает главным образом при протекании ионных токов через невозбудимые (не реагирующие на сигналы) каналы, избирательно проницаемые для ионов. Так например, особые K+-каналы – «калиевые каналы утечки», открыты даже в состоянии покоя. К пассивному ответу способны все клетки любых тканей. Это основа раздражимости, но не возбудимости.
2. Активные электрические ответы, характерные только для возбудимых тканей (нервов, мышц, рецепторов), обусловлены открыванием и закрыванием множества мельчайших ионных каналов в ответ на раздражение. Поведение совокупности каналов, проницаемых для того или иного иона, определяет перемещение этого иона в том или ином направлении в зависимости от соответствующего электрохимического градиента.
Срабатывание одних каналов зависит от связывания с особыми рецепторными участками мембраны молекул медиаторов или мессенджеров (рецептор-зависимые каналы), других – от воздействия специфических раздражителей (свет, температура и т. д.), третьих – от изменения мембранного потенциала (потенциал-зависимые каналы, коротко – φ-зависимые). Последние особенно важны: только их наличие обеспечивает развитие распространяющегося возбуждения (см. далее).
Большинство возбудимых каналов в той или иной степени обладает избирательной проводимостью (селективностью): они пропускают какой-либо ион (или группу ионов) лучше, чем остальные ионы. Поэтому каналы часто называют в соответствии с тем, для каких ионов они в норме проницаемы: так, натриевые каналы могут пропускать не только натрий, но также и литий, однако в норме при возникновении нервных импульсов через эти каналы проходит именно натрий. При открывании одного канала через мембрану протекает лишь слабый ток, т.к. проходит мало ионов. Однако при одновременном срабатывании множества подобных каналов через мембрану идет весьма значительный ток, вызывающий заметное изменение мембранного потенциала.
Подводя предварительные итоги, мы можем выделить три процесса, необходимые для возникновения активных электрических сигналов (т. е. возбуждения мембраны):
1. Активный перенос ионов мембранными насосами, в результате которого по разные стороны мембраны создается неравномерное распределение различных ионов.
2. Наличие электрохимического градиента, обусловленного неравномерным распределением различных ионов.
3. Открывание ионных каналов, избирательно проницаемых для того или иного иона. Через открытые каналы может течь ионный ток, движущей силой для которого служит электрохимический градиент для данного иона. (к оглавлению)
Возбуждение может быть локальным (местным) и распространяющимся.
Локальное возбуждение (рецепторный потенциал – РП) захватывает лишь отдельные структуры возбудимых образований. Оно характеризуется следующими особенностями:
а) возникает в участках клеточных мембран, специализирующихся на восприятии внешних раздражителей,
б) не распространяется по мембране,
в) до определенной степени, пропорционально силе и длительности воздействия раздражителя, т.е. нарастает по мере увеличения раздражения.
Это объясняется тем, что развитие локального возбуждение связано с работой каналов первых двух типов, но не φ-зависимых каналов. Каналы открываются лишь на том участке, на который воздействовал раздражитель (нет распространения). Количество открытых каналов (а значит и количество входящих в клетку катионов и степень изменения мембранного потенциала) пропорционально интенсивности воздействия раздражителя (3 молекулы медиатора откроют 3 канала, а 20 – 20 каналов и мембранный потенциал изменится значительнее). Оговорка «до определенной степени» связана с ограниченным количеством каналов, и если они на данном участке открыты все, то дальнейшее увеличение силы воздействия раздражителя ничего не изменит.
В возбудимых тканях относительно низкой организации (железистый эпителий) локальное возбуждение, приводящее к специфической деятельности клеток, является единственной формой ответа на раздражение. Это связано с отсутствием в мембранах клеток железистого эпителия φ-зависимых каналов.
Потенциал действия
Ключевые понятия:
- ПП (потенциал покоя), локальное возбуждение (РП), потенциал-зависимые каналы (см. выше);
- ПД (потенциал действия) – распространяющееся возбуждение;
- деполяризация мембраны – изменение мембранного потенциала на менее отрицательный и даже положительный;
- ПорП (пороговый потенциал) – значение деполяризации, при котором реагируют и открываются φ-зависимые каналы;
- овершут – пИковое превышение мембранным потенциалом нулевого значения (положительный заряд);
- реполяризация – понижение мембранного потенциала после деполяризации до значения ПП;
- гиперполяризация мембраны – период, когда мембранный потенциал имеет значения ниже ПП;
- рефрактерность – невосприимчивость мембраны к новым воздействиям (может быть абсолютной и относительной).
- супернормальность - повышенная возбудимость мембраны.
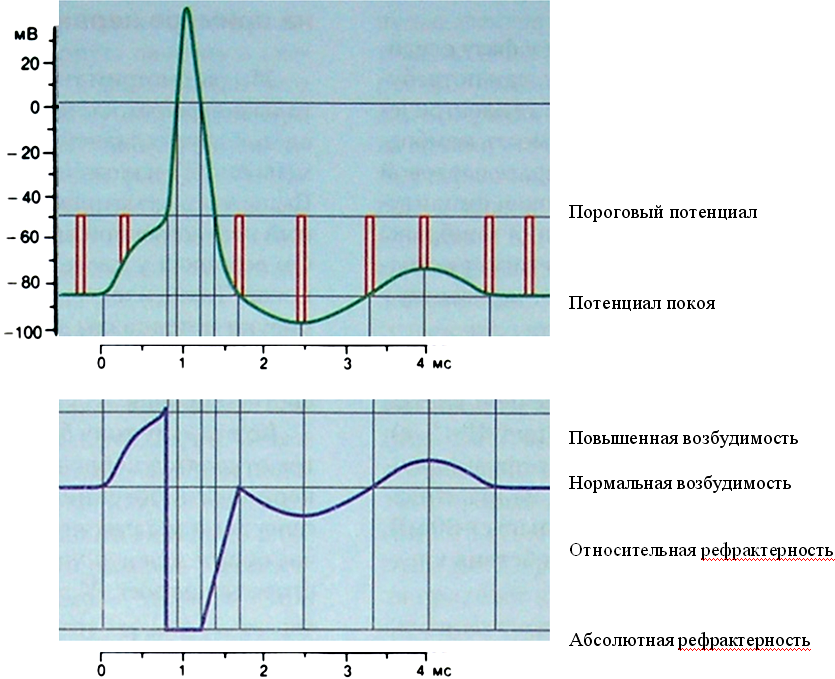
Механизм развития. В клетках нервной и мышечной ткани, имеющих потенциал-зависимые каналы, возбуждение развивается следующим образом (мВ – милливольты, мс – миллисекунды; отсчет времени начинается с момента воздействия раздражителя):
1-ый этап (0 – 0,7мс) – развитие локального возбуждения (см. выше).
2
 -ой
этап – если изменение мембранного
потенциала из-за локального возбуждения
достигает определенной пороговой
величины, т.е. достигнут ПорП
(0,7-ая мс), то открываются ВСЕ
φ-зависимые каналы, и
-ой
этап – если изменение мембранного
потенциала из-за локального возбуждения
достигает определенной пороговой
величины, т.е. достигнут ПорП
(0,7-ая мс), то открываются ВСЕ
φ-зависимые каналы, и
3-ий этап (0,7 – 1,7мс) – мгновенно локальное возбуждение переходит в распространяющееся (Потенциал действия – ПД) т.к. изменение мембранного потенциала одного участка, приводит к открытию φ-зависимых каналов на соседнем, и т.д. ПД развивается по принципу «все или ничего» т.е. сразу приобретает максимальную амплитуду (открыты ВСЕ φ-зависимые каналы) и достигает на овершуте значения +40мВ.
4-ый этап – следовые потенциалы – гиперполяризационный (1,7 – 3,3мс) и деполяризационный (3,3 – 5мс).
Таким образом, локальное возбуждение в нервной и мышечной тканях можно рассматривать как начальный этап, пусковой механизм распространяющегося возбуждения.
Мерой возбудимости тканей является порог возбудимости (на графике он показан красной двойной линией). Это минимальная сила раздражителя (действующего внезапно и достаточно длительно), которая способна вызвать процесс возбуждения. Раздражители меньшей или большей силы называются соответственно подпороговыми и сверхпороговыми. Величина порога характеризует сравнительную возбудимость разных тканей. Чем ниже порог возбуждения, тем выше возбудимость, т.е. тем меньшая сила стимула требуется для достижения ПП. Иными словами: если мембрана уже частично деполяризована, то ПорП достигается под действием раздражителя меньшей силы, а, если гиперполяризована – все наоборот. Порог возбуждения нерва ниже, чем порог возбуждения мышцы и особенно железы. Функциональное состояние ткани (работа, утомление, уровень метаболизма) также влияют на величину порога. (к оглавлению)
Н а нижнем графике рисунка показано изменение возбудимости мембраны в момент развития ПД. Проанализируйте его и сопоставьте изменение возбудимости мембраны с этапами развития ПД (верхний график).
Для того, чтобы понять причины особенностей ПД и возбудимости, необходимо разобраться с механизмом работы φ-зависимых каналов.
Механизм работы φ-зависимых каналов.
Процессы, приводящие к открыванию или закрыванию таких каналов, называются воротными. Представим себе типичную возбудимую клетку в состоянии покоя. Пусть ее мембранный потенциал покоя (ПП) составляет –75 мВ, а пороговый потенциал (ПорП) –25мВ. Следовательно, деполяризация на 50 мВ (с –75 до – 25 мВ) приведет к активации натриевых каналов, расположенных в мембране. Каналы представляют собой молекулы белка, вкрапленные в мембранный фосфолипидный бислой. Значит, при деполяризации на 50 мВ в этом бислое и в канальных белках возникает изменение электрического напряжения. Полярные группировки белков–каналов «чувствуют» такие изменения по электростатическому принципу «притяжение-отталкивание» и отвечают на них конформационными перестройками (изменением формы) каких–либо участков белковых молекул приводящим к перекрыванию или открыванию внутренней полости канала. Эти конформационные перестройки и лежат в основе воротных процессов, управляющих электровозбудимыми каналами. В клетках обнаружены воротные механизмы работы натриевых, калиевых, кальциевых и т.п. каналов. Рассмотрим воротные процессы на примере натриевых электровозбудимых каналов.
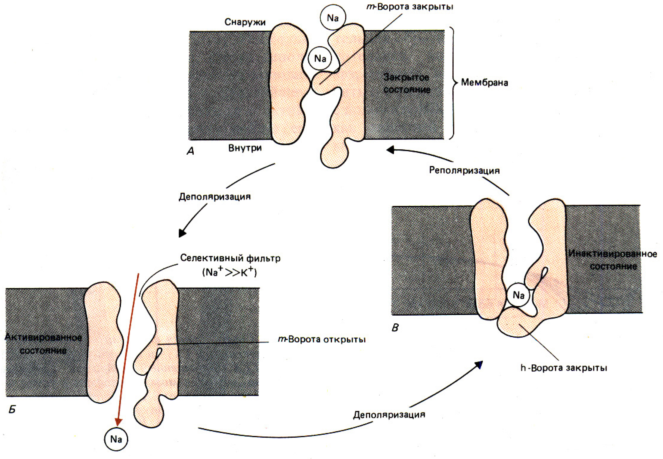
Натриевые каналы представляют собой крупные интегральные канальные белки с двумя участками (m–ворота и h–вopoтa). Эти «ворота» способны при изменении своего положения перекрывать или открывать полость канала.
О
 сновные
состояния натриевых каналов:
сновные
состояния натриевых каналов:
1) В покое: m–ворота (активационные) закрыты, h–вopoтa (инактивационные) открыты; канал не пропускает ионы Na+; значение мембранного потенциала соответствует ПП. (рис.А)
2) При деполяризации до значения ПорП m–ворота открываются, канал активируется и начинает пропускать ионы Na +.(рис.Б)
3) На пике овершута (+40мВ) закрываются h–вopoтa, и канал инактивируется и начинается реполяризация мембраны. (рис.В)
4) Реполяризация до уровня потенциала покоя приводит к закрыванию m–ворот; h–вopoта пока закрыты, но в этом состоянии канал можно активировать сверхпороговым деполяризующим стимулом.
5) Следовая гиперполяризация мембраны приводит к открытию h–вopoт; мембранный потенциал постепенно возвращается к ПП; канал находится в исходном состоянии (рис.А).
Обратите внимание, что на этапе, когда закрыты и m–ворота, и h–ворота, между ними остается «заперто» некоторое количество ионов Na+. При открывание h–ворот эти ионы устремляются в клетку, вызывая следовую деполяризацию мембраны на 3,3 – 5мс после нанесения раздражения (см. график ПД). В этот период, поскольку мембрана уже частично деполяризована, для достижений ПорП ее надо перезарядить на меньшее значение, и даже воздействие подпорогового раздражителя может привести к достижению значения ПорП, а значит вызвать ПД. Это явление повышенной возбудимости мембраны называется «супернормальность».
В развитии ПД, помимо натриевых каналов принимают участие и калиевые электровозбудимые каналы. Они открываются на пике овершута в дополнение к K+-каналам утечки, открытым в состоянии покоя и обеспечивают быструю реполяризацию мембраны.
(к оглавлению)