

Первичные радикалы (радикалы кислорода, окись азота, радикал коэнзима q). Первичные радикалы
Основные радикалы, образующиеся в клетках - это радикалы кислорода (супероксид и гидроксильный радикал), монооксид азота, радикалы ненасыщенных жирных кислот, радикалы, образующиеся в окислительно-восстановительных реакциях (например, убихинол). Радикалы образуются также при действии ультрафиолетовых лучей и в ходе метаболизма некоторых чужеродных соединений (ксенобиотиков), в том числе некоторых препаратов, одно время применявшихся в качестве лекарств.
Активные формы кислорода
Основная масса молекулярного кислорода, потребляемого клетками нашего организма, непосредственно восстанавливается до воды, окисляя органические субстраты в цепях переноса электронов. Меньшая часть кислорода расходуется на неполное окисление органических соединений. Наконец, заметная часть кислорода восстанавливается клетками нашего организма до супероксидного радикала. Так клетки – фагоциты (моноциты и гранулоциты крови и тканевые макрофаги) выделяют супероксид в реакции, катализируемой ферментным комплексом – НАДФН-оксидазой:
НАДФН + 2O2 -> НАДФ+ + 2O•- (супероксид) |
Дальнейшая судьба супероксидных радикалов может быть разной (см. схему на рис.2). В норме и при отсутствие ионов металлов переменной валентности супероксидные радикалы превращаются в перекись водорода; эта реакция катализируется ферментом супероксиддисмутазой (СОД) (реакция 2 на рис.2):
2O•- -> H2O2 + O2 |
Клетки-фагоциты используют перекись водорода, превращая ее в гипохлорит – соединение, разрушающее стенки бактериальных клеток; эта реакция катализируется ферментом миелопероксидазой (реакция 3 нарис.2):
H2O2 + Cl- -> H2O + ClO- |
Избыток перекиси водорода удаляется под действием двух ферментов: глутатионпероксидазы или каталазы (реакция 4 на рис.2):
|
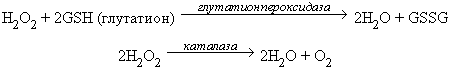
В условиях патологии могут произойти нарушения либо системы защитных ферментов (в частности, снижение активности СОД), либо ферментных систем, связывающих ионы железа в плазме крови (церулоплазмин и трансферрин) и в клетках (ферритин). В этом случае супероксидные радикалы и перекись водорода вступают в альтернативные реакции: Образование двухвалентного железа из трехвалентного (реакция 7 на рис.2):
Fe3+ + O•- -> Fe2++ O2 |
Реакция перекиси водорода и гипохлорита с ионами двухвалентного железа (реакции 9 и 10 на рис.2):
Fe2+ + H2O2 -> Fe3+ + HO- + HO· (радикал гидроксила) |
Fe2+ + ClO- + H+ -> Fe3+ + Cl- + HO· (радикал гидроксила) |
Совокупность продуктов, образуемых активированными клетками-фагоцитами (радикалы супероксида и гидроксила, перекись водорода и гипохлорит) называют активными формами кислорода; некоторые авторы называют гипохлорит и продукты его метаболизма в тканях (такие как хлорамины R-NHCl) называют активными формами хлора. Радикалы гидроксила химически исключительно активны и вызывают повреждение белков, нуклеиновых кислот и липидов биологических мембран. Особенно тяжелые последствия имеют две последние реакции. Радикалы ·OH вызывают разрыв нитей ДНК, обладая, в зависимости от ситуации, мутагенным, канцерогенным или цитостатическим действием. С другой стороны, реагируя с ненасыщенными жирными кислотами, входящими в состав мембранных липидов, радикалы гидроксила инициируют цепную реакцию их пероксидации (перекисного окисления).