

19. Перенесення електронів в анаеробних умовах (анаеробне дихання)
У результаті фосфорилювання, спряженого з перенесенням електронів у процесі дихання аеробних мікроорганізмів, утворюється набагато більше АТФ, ніж при субстратному фосфорлюванні, яке відбувається під час бродіння. Тому не дивно, що в процесі еволюції виник і зберігся такий тип метаболізму, у якому водень від органічного субстрату переноситься на "зв'язаний" кисень (нітрат, сульфат, карбонат ти ін.); ці сполуки відновлюються воднем субстрату. Здатність переносити електрони на такі сполуки дає бактеріям змогу окиснювати субстрати без участі молекулярного кисню і одержувати більше енергії, ніж це можливо у процесі бродіння.
Такі бактерії мають систему транспорту електронів, містять цитохроми. Одержання енергії фосфорилюванням, спряженим з перенесенням електронів на такі термінальні акцептори, як нітрат, сульфат тощо, називають анаеробним диханням. Залежно від природи неорганічного акцептора водню розрізняють нітратне, сульфатне, карбонатне та інше дихання (рис. 27.1).
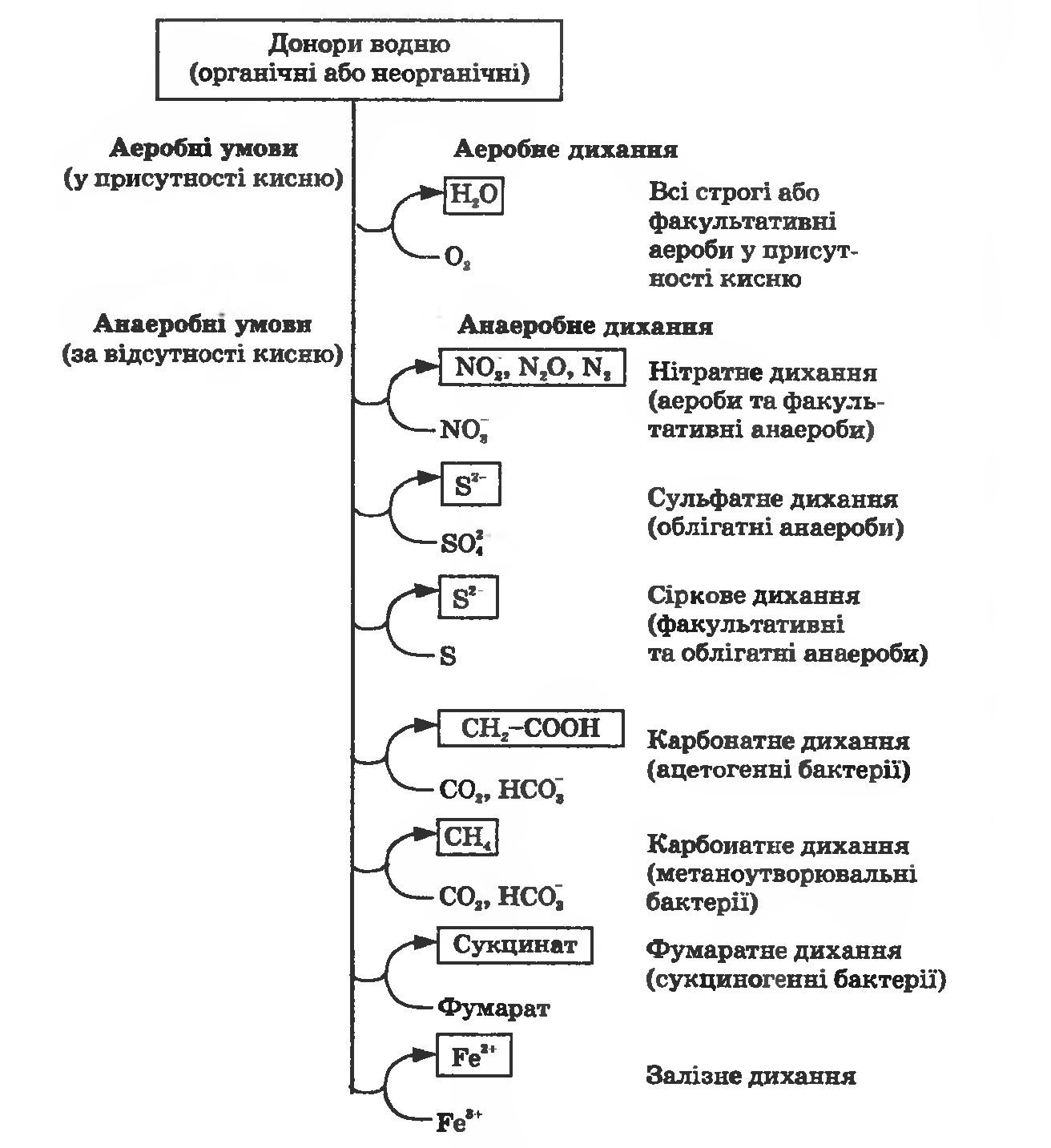
Рисунок 27.1. Процеси одержання енергії окиснювальним фосфорилюванням в аеробних та анаеробних умовах (аеробне та анаеробне дихання)
27.1Денітрифікауія та відновлення нітрату
Мікроорганізми використовують нітрат:
1) для синтезу азотвмісних клітинних компонентів. Така асиміляційна нітратредукція може відбуватися як в аеробних, так і в анаеробних умовах;
2) для нітратного дихання (дисиміляційна нітратредукція), при цьому нітрат в анаеробних умовах є термінальним акцептором водню (електронів).
В обох цих випадках нітрат спочатку відновлюється до нітриту за допомогою нітратредуктази. У процесі асиміляційної нітратредукції утворений нітрит відновлюється до аміаку за допомогою нітритредуктази, на що витрачається шість електронів. Електрони надходять від НАД(Ф)Н (у грибів і бактерій) або фередоксину (у рослин і деяких бактерій). Аміак використовується для синтезу клітинних компонентів. Денітрифікуючі бактерії здатні відновлювати нітрат через нітрит до газоподібного закису азоту (N2О) та азоту (N2). Цей процес денітрифікації виявлений тільки у факультативних аеробів. Деякі денітрифікатори можуть рости, використовуючи як термінальний акцептор водню не тільки нітрат, а й нітрит, а також іноді й закис азоту. Представниками денітрифікуючих бактерій є ґрунтові бактерії, наприклад, бацили, Paracoccus denitrificans.
27.2. Утворення сірководню у процесі відновлення сульфату
Сульфатредукуючі (сульфатовідновлювальні) бактерії характеризуються здатністю до перенесення водню на сульфат як термінальний акцептор електронів і таким чином до відновлення сульфату до сульфіду. Цей процес називається сульфатним диханням, або дисиміляційною сульфатредукцією. Основним продуктом цього процесу є сірководень. Більша частина сірководню, який є в природі, утворюється завдяки життєдіяльності бактерій-сульфатредукторів.
Сульфатредукуючі бактерії — це фізіологічна група, для якої характерним є здатність до утворення сірководню з сульфату. Сульфатовідновлювальні бактерії, на відміну від нітратредукторів, є облігатними анаеробами. Донорами водню служать прості низькомолекулярні сполуки, які утворюються внаслідок анаеробного розкладання біомаси (в основному целюлози): лактат, ацетат, пропіонат, бутират, форміат, етанол, вищі жирні кислоти, молекулярний водень. За ступенем засвоєння органічних кислот розрізняють дві групи сульфатредукторів:
1) окиснюють донор не повністю та виділяють оцтову кислоту. Такими є представники спороутворювального роду Desulfotomaculum (наприклад, D. nigrificans) та неспороутворювального роду Desulfovibrio (D. vulgaris, D. desulfuricans). Ці бактерії не мають повного циклу трикарбонових кислот;
2) частина представників цієї групи може рости, використовуючи спирти, ацетат, вищі жирні кислоти або бензоат, тобто є хемоорганогетеротрофами, а інші здатні навіть до хемолітоавтотрофного росту у присутності водню та форміату. Деякі штами здатні синтезувати клітинні компоненти з ацетату, якщо донором водню служить Н2, тобто є хемолітогетеротрофами. До них належать спороутворювальні (Desulfotomaculum acetoxidans) та неспороутворювальні палички (Desulfobacter), коки (Desulfococcus), сарцини (Desulfosarcina), нитчасті форми, які рухаються ковзанням (Desulfonema), та ін.
Деякі сульфатредуктори можуть рости у присутності Н2 і сульфату як єдиних джерел енергії. Здатність же до відновлення сульфату за допомогою молекулярного водню і утворення при цьому великих кількостей сірководню, не пов'язаного з помітним ростом, є характерною для більшості сульфатредукторів.
Асиміляційна та дисиміляційна сульфатредукція. Майже всі бактерії, гриби та зелені рослини здатні використовувати як джерело сірки сульфат. Вони одержують із сульфату сульфід, необхідний для синтезу сірковмісних амінокислот, шляхом асиміляційної сульфатредукції (рис. 27.2). Сульфат двічі активується за рахунок АТФ з утворенням аденозин-5-фосфосульфату (АФС) та фосфоаденозин-5-фосфосульфату (ФАФС) (ферменти АТФ-сульфурилаза, АФС-кіназа). Лише цей двічі активований сульфат може відновлюватись спочатку до сульфіту (ФАФС-редуктаза), а потім до сульфіду (сульфітредуктаза).
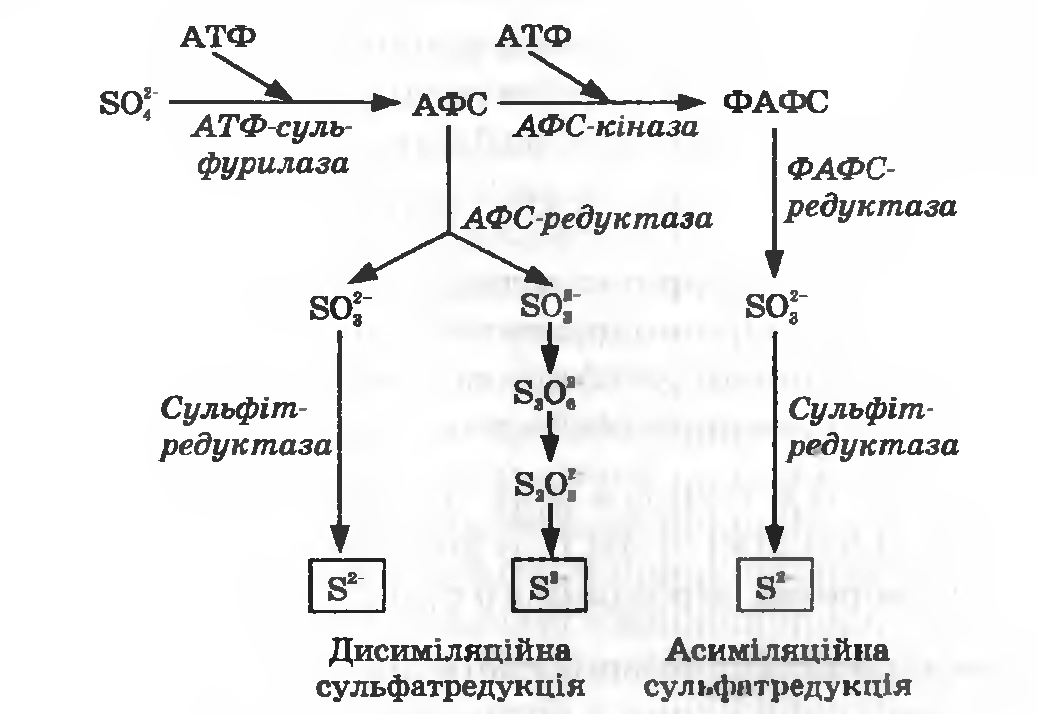
Рисунок 27.2. Схема дисиміляційіюго та асиміляційного відновлення сульфату
Перший етап дисиміляційної сульфатредукції є аналогічним процесу асиміляції сульфату. Далі АФС відновлюється (фермент АФС редуктаза), причому відновлення може відбуватися двома шляхами:
1) за допомогою сульфітредуктази сульфат прямо (без утворення проміжних продуктів) відновлюється до сульфіду, на що витрачається шість електронів;
2) послідовне триетапне відновлення з утворенням проміжних продуктів таких, як тритіонат і тіосульфіт.
Бактерія Desulfuromonas acetoxidans здатна використовувати елементарну сірку як акцептор водню у процесі анаеробного перенесення електронів. Сірка при цьому відновлюється до сірководню. Ця виділена з морської води бактерія здатна повністю (до СО2 та води) окиснювати етанол та ацетат.
Поширення та роль сульфатредукторів у природі. Сульфатредуктори зустрічаються в основному в сірководневому мулі, де органічні речовини піддаються анаеробному розкладанню. Основну масу сірководню, утворюваного в природі, слід вважати кінцевим продуктом сульфатного дихання. Більшість сіркових родовищ мають невулканічне походження. Це так звана біогенна сірка, яка утворилась шляхом відновлення сульфатів морської води за допомогою сульфатредукуючих бактерій.