

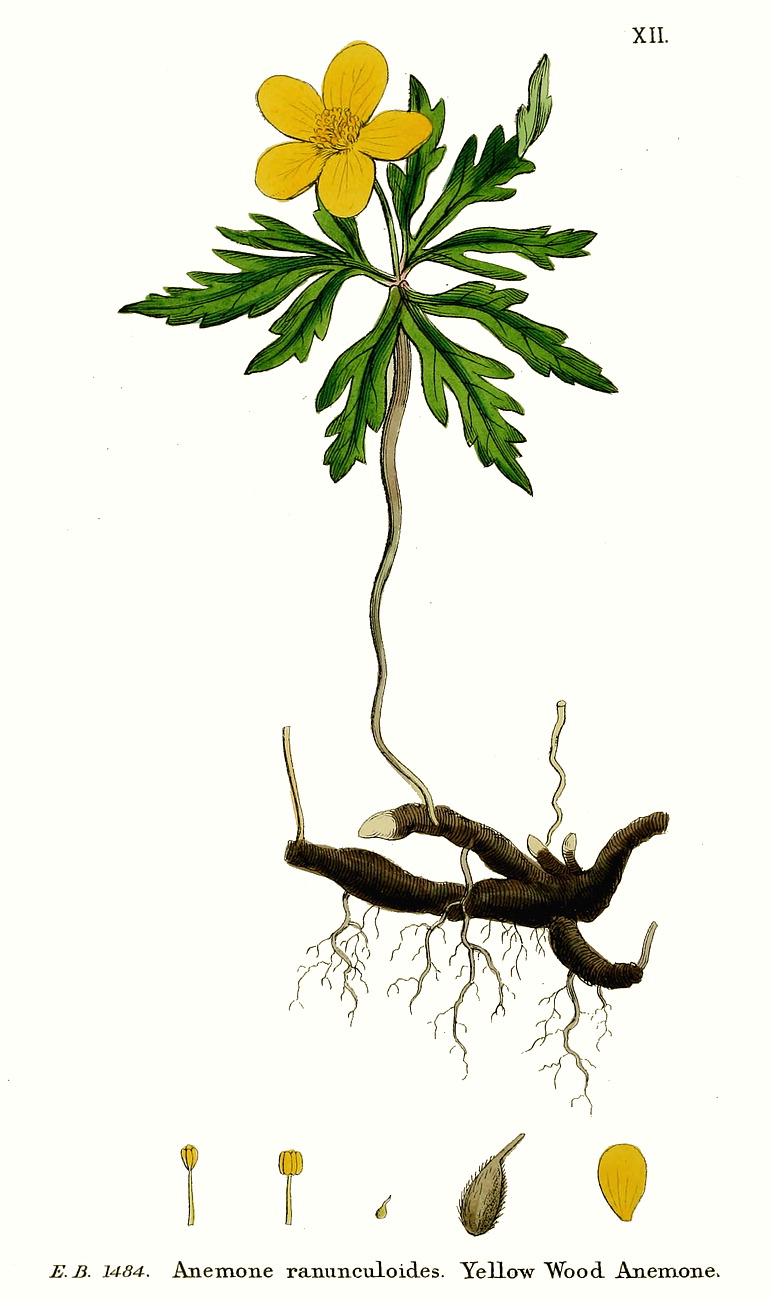
2.3 Ветреница лютичная (Anemone ranunculoides l.)

Многолетнее травянистое растение с подземным (гипогеогенным) ползучим, обычно разветвленным корневищем. Надземная часть представлена длинночерешковыми зелеными листьями вегетативных побегов и цветоносами с горизонтально распростертыми: листьями покрывала. Корневище мясистое, несет широкие короткие чешуи, охватывающие каждая до 1/2 его периметра и представляющие собой основания влагалищных листьев, отгибы которых быстро опадают. Чешуи не закрывают пазушных почек; последние постепенно углубляются в мякоть корневища. Корневище почти: полностью лишено механической ткани (иногда слабо развита колленхима во внешней коре); проводящие пучки закрытого типа, без механических обкладок; утолщение корневища идет за счет разрастания основной паренхимы. Последнее междоузлие каждого годичного прироста корневища тоньше остальных, поэтому многолетние его ветви четковидные и при механических повреждениях легко распадаются на отдельные «членики» (участки годичного прироста).
Основные (скелетные) ветви корневища у взрослого растения нарастают симподиально: побеги каждого следующего порядка образуются из пазушных почек низовых листьев цветоноса и, трогаясь в рост одновременно с цветением, формируют в течение вегетации плагиотропную часть («членик») корневища; на ее верхушке закладывается почка возобновления и весной вытягивается ортотропная часть побега — цветонос, полностью отмирающий после плодоношения. Таким образом, корневище складывается из монокарпических побегов, развивающихся по моноциклическому типу (Серебряков, 1952). Нередко от основания цветоноса отходят два (изредка три) новых побега (верхушечное ветвление); на более старых участках корневища из спящих почек возникают боковые ветви (боковое ветвление), которые обычно не сразу переходят к цветению и некоторое время нарастают моноподиально. Ветвление после отмирания коммуникационных участков корневища обеспечивает вегетативное размножение.
Нецветущие ветви корневища несут по одному зеленому листу с длинным, иногда скрученным, черешком; пластинка тройчато или пальчаторассеченная, средняя доля трех-, боковые двух-четырехраздельные. Листья покрывала всегда тройчаторассеченные, почти сидячие, доли их ромбические, неравнобокие, с крупными заостренными зубцами; все покрывало горизонтально распростертое. Анатомически листья отличаются слабым развитием столбчатого мезофилла и механической ткани, невысоким содержанием хлорофилла и небольшой общей протяженностью жилок (Карпенко, 1964, 1968а; Горышина, 1969).
Цветок 1,5—3 см в диаметре, один на конечном цветоносе или их 2—3 (редко больше) на цветоножках, пазушных по отношению к листьям покрывала; 5 (изредка 6—7) желтых листочков околоцветника покрыты с внешней стороны тонкими прижатыми волосками. Тычинки в 3 раза короче листочков околоцветника. Пестики с короткими изогнутыми столбиками, завязи и плоды опушенные. Фертильная семяпочка одна, с одним интегументом, крассинуцеллятная, с удлиненным фуникулюсом, перемещающим ее к моменту оплодотворения в гипертропное положение; вероятно, закладывается семяпочка как атропная (Иванова, 1966). Зародышевый мешок моноспорический, Polygonatum-типа, с очень крупными клеточными элементами (Иоффе, 1958).
Плодики около 3 мм длины, с коротким носиком, опушенные, распространяются муравьями (Hegi, 1935; Трофимов, 1939; Рысина, 1969). Семена без перисперма. Ко времени опадения плодиков зародыш в семени не дифференцирован и представляет собой маленькое шаровидное тельце, лежащее близ носика орешка. Развитие зародыша идет после опадения плодика и к осени того же года семя становится способным к прорастанию (Thome, 1886; Goebel, 1933; Серебряков, 1952; Рысина, 1973); в это время зародыш имеет две крупные полусферические семядоли, зародышевый корешок и зачаток первого листа; эндосперм полностью расходуется.
Диплоидный набор хромосом 32 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Ареал вида европейско-малоазиатский. Растение встречается во всей Европе (отсутствует в Великобритании, на севере Скандинавии, на юге Италии и Греции). В СССР обычно в лесах европейской части, в дубравах лесостепи, лесах Крыма, Предкавказья, заходит на Кавказ и в Малую Азию; на востоке доходит до Урала, где сменяется A. uralensis Fisch.; близкими видами являются также сибирские A. coerulea DC., A. jenissensis (Korsh.) Kryl. и дальневосточная A. debilis Fisch. Шарфеттер (Scharfetter, 1953) рассматривает эти виды как производные одного плиоценового (типа keiskeana), мигрировавшие из горных областей с.-з. Китая на восток (в Корею, Японию и Америку) и на север и запад — по горам Алтая, Сибири, Туркмении на Урал, в Карпаты, Альпы и до Пиренеев, расселившиеся затем в равнинных лесах. На протяжении всего ареала ветреница лютичная встречается в буковых, дубово-буковых, дубово-грабовых, дубово-ясеневых и липовых насаждениях, заходя в елово-широколиственные. Хорошо выносит осветление, сохраняется во вторичных сообществах (осинники, редколесья, кустарниковые заросли), но не выдерживает уплотнения и задернения почвы (Карписонова, 1961, 1967). В чистых вторичных березняках, а также в участках елово-широколиственных лесов при временном господстве ели и разрастании зеленых мхов быстро исчезает. В нарушенных дубравах паркового типа сохраняется возле отдельных кустов и стволов, в западинках микрорельефа и в оврагах, где больше скапливается опада и не уплотняется почва.
В Московской обл. ветреница лютичная распространена повсеместно в дубравах и дубо-липняках, кустарниковых зарослях с господством лещины; нередко встречается в смешанных лесах, тяготея к ценозам неморальной флоры, а также в сероольшаниках.
Онтогенез
Онтогенез вида (жизненный цикл) изучен довольно полно (Irmisch, 1856; Голубев, 1956; Смирнова, 1968; Старостенкова, 1971; Рысина, 1973; Гуланян, 1974). Плодики ветреницы лютичной опадают в конце мая — начале июня; дозревание семян в них идет в течение 2,5—4 месяцев. Часть плодиков прорастает осенью в год образования, часть — следующей весной; плодики могут сохраняться жизнеспособными и не прорастать в течение ряда последующих лет (продолжительность сохранения жизнеспособности неизвестна).
Прорастание подземное. Плодовую оболочку пробивает сначала корешок, затем появляется удлиненный гипокотиль; семядоли же и зачаток первого зеленого листа остаются в оболочке плода. При осеннем прорастании первый лист не развертывается — он появляется над землей весной. Длинный, тонкий, нередко извитой черешок этого листа несет тройчатонадрезанную или тройчаторассеченную пластинку длиной 1—1,5 см; средняя доля ее цельная. Гипокотиль клубневидно утолщается, хорошо заметен главный корень. Семядоли сохраняются в плодовой оболочке до конца вегетации первого года жизни сеянца. К этому времени у него есть один-два боковых корешка, ось над гипокотилем несколько вытянута, сформирована почка возобновления. Осенью выше гипокотиля появляется придаточный контрактильный корень, придающий оси горизонтальное положение. В течение первых трех-четырех лет жизни ювенильного растения облик его мало меняется: сохраняется главный корень, заметен гипокотиль — тот и другой отмирают на 4—5-й год жизни; ежегодно образующийся единственный зеленый лист имеет тройчатую пластинку с почти цельной средней и слабо надрезанными боковыми долями; годичные приросты моноподиальной оси незначительны (образуется 1—3 узла). Особи с отмершим гипокотилем могут быть названы имматурными: листовые пластинки у них обычно имеют пять долей (боковые доли тройчатого листа до основания рассечены), средняя доля глубоко надрезанная, но без заостренных зубцов; годичные приросты корневища достигают 0,9—1,2 см.
Взрослые вегетативные растения сохраняют моноподиальное нарастание осей, листья более крупные, средняя доля тройчато раздельная, с зубчатым краем, годичный прирост корневища — до 3 см, обнаруживается боковое ветвление. После отмирания старых участков корневища (через 5—9 лет после их образования) ветви обособляются, начинается вегетативное размножение; боковые ветви до отделения не ветвятся, часть из них очень мелкие с незначительным годичным приростом и ювенильного типа листьями; отделяясь от материнской особи в заросли взрослых растений они, вероятно, не имеют возможности дальнейшего развития и образуют «отпад».
Период догенеративной вегетации у ветреницы лютичной в естественных условиях оказывается довольно продолжительным и длится не менее 10 лет (иногда, вероятно, значительно дольше). В условиях культуры он укорачивается — растение может зацветать на втором-третьем году жизни (Евтюхова, 1968).
С переходом в генеративное состояние на верхушках основных ветвей корневища закладываются цветоносы, нарастание этих ветвей становится симподиальным, начинается верхушечное ветвление, более интенсивным становится и боковое ветвление, годичный прирост корневищ достигает нескольких сантиметров, увеличивается число придаточных корней на каждом «членике», усиливается контрактильная деятельность корней, корневище несколько заглубляется. При отмирании старых участков корневищ у таких растений освобождаются уже разветвленные корневищные системы. Распад корневищ ускоряется тем, что при отмирании крупных цветоносов некротические процессы по мягким тканям распространяются на подземные части, на верхушке плагиотропного побега прошлого года появляется изъязвление с раневой пробкой, а впоследствии корневище в этом месте разламывается: отделяются молодые (с одним-двумя годичными приростами и верхушечными почками) участки корневища, жизнеспособной остается и старая его часть, лишенная верхушки, но с боковыми ветвями, усиливающими рост. Генеративные особи, таким образом, обладают наибольшей вегетативной подвижностью, обеспечивают наиболее интенсивное вегетативное размножение, семенное возобновление, а также высокую насыщенность побегами занятого растением пространства.
В начале генеративного периода (молодые генеративные растения) оси корневищ сохраняют в основном моноподиальный характер, так как цветение и последующее образование симподиев происходят не ежегодно, верхушечного ветвления почти нет. С увеличением мощности особей (средневозрастные генеративные растения) цветение и симподиальное нарастание, а также верхушечное ветвление становятся ежегодными, образуется много боковых ветвей, которые скоро (на второй-третий год) достигают цветения. Старение особей приводит к уменьшению годичных приростов корневища, а следовательно, вегетативной подвижности; уменьшается верхушечное ветвление, цветение основных ветвей идет с перерывами, вновь возникающие боковые ветви мелкие (признаки стареющих генеративных растений). В этом возрастном состоянии образуется много «отпада», в значительной своей части, вероятно, погибающего.
Старые вегетативные особи сохраняют крупные скелетные ветви корневища, но с моноподиальным нарастанием или вообще лишенные верхушки; прирост незначителен. Сенильное состояние характеризуется прекращением деятельности точек роста скелетных ветвей; постепенно корневище распадается на короткие участки, несущие почти не нарастающие в длину веточки с истощенными точками роста. Жизнь таких особей может продолжаться пока на старых приростах корневища сохраняются живые почки, образующие новые боковые веточки с маленькими зелеными листьями. В этом возрастном состоянии почти нет прироста, вегетативное размножение отсутствует, подвижность растения ничтожна.
Участки (членики) корневища ветреницы лютичной нередко обнаруживаются в почве живыми, но без боковых веточек с листьями. Вероятно, отделившись при механическом повреждении или естественном распаде корневища, они могут некоторое время сохраняться в состоянии вторичного покоя. Условия, продолжительность покоя и роль таких особей в жизни природных популяций, по-видимому, не изучены.
Общая продолжительность жизненного цикла ветреницы лютичной (от прорастания семени до полного отмирания особи) в природных популяциях, по мнению О. В. Смирновой (1968), составляет не менее 40—50 лет, а при благоприятных для вегетативного размножения условиях—в несколько раз больше за счет жизни омоложенного вегетативного потомства.
Сезонный ритм развития
Сезонный ритм развития ветреницы лютичной изучен довольно подробно (Проскоряков, 1926; Кожевников., 1929, 1931; Трофимов, 1939; 1954; Серебряков, 1947; Шик, 1953; Скрипчинокий, Скрипчинский, 1961; Смирнова, 1968; Горышина, 1969; Старостейкова, 1971). Как и у некоторых других растений широколиственного леса, начало видимого роста отмечается еще под снегом; развертывание листьев и цветение начинаются сразу после таяния в лесу основной массы снега и оттаивания почвы. Надземная вегетация продолжается от 3 до 7 недель. Наименьшая продолжительность вегетации отмечается в дубравах лесостепи (Горышина, 1969); она возрастает по мере продвижения на запад (Мельниченко, 1934) и на север (Трофимов, 1939, 1949).
Цветки появляются почти одновременно с развертыванием листьев. Цветение продолжается одну-две недели. Цветки открываются в солнечную или достаточно теплую погоду, а на ночь, во время дождя или заморозка остаются закрытыми. Отдельный цветок продолжает открываться от 3—4 до 7—10 дней. Опыление производят различные насекомые, собирающие пыльцу; цветки, вероятно, протерогиничны (Hegi, 1935). Оплодотворение протекает по обычному для покрытосеменных типу (Иоффе, 1958). Созревание и опадение плодиков происходит за 7-—10 дней до завядания надземных частей.
Рост листьев и вытягивание цветоносов продолжаются во время цветения и после него до самого конца вегетации и полного завядания (Горышина, 1969). Одновременно с ростом надземных частей идет нарастание корневищ. Отмирание надземных частей начинается с потери ими тургора, завядания и полегания. Дальнейшее их разрушение идет очень быстро при высокой температуре и солнечном освещении, но может значительно задерживаться в оврагах и под густым покровом листьев других растений. Вскоре после отмирания надземных побегов прекращается рост корневищ, очень замедлены в это время процессы формообразования в почках возобновления.
В конце лета наступает активизация роста — быстро идет формирование зачатков листьев и цветков в почках, появляются придаточные корни на молодых приростах корневища. Ко времени осенних холодов полностью сформированы в почках побеги будущего года и развиты корни, которые обеспечат начало вегетации весной. Конечные междоузлия на молодых приростах корневища вытягиваются и почки возобновления поднимаются к поверхности почвы. Поздней осенью до наступления устойчивых морозов идут процессы споро- и гаметогенеза в цветках (Шик, 1953). Часто осенью цветочные почки раскрываются, появляются маленькие зеленоватые пластинки низовых листьев, зеленеют листья покрывала, одевающие зачаток цветка. При раннем наступлении осенних холодов почки не раскрываются. Весной низовые листья цветоноса имеют вид чешуй и не несут зеленых пластинок. По наблюдениям Т. К. Горышиной (1969), осенее зеленение не имеет значения для накопления ассимилятов. В предвесеннее время под снегом могут увеличиваться в размерах и вытягиваться цветоносы, идет образование хлорофилла. Однако в малоснежные зимы и при сильном промерзании почвы подснежный рост и зеленение отсутствуют, на дальнейшем развитии это существенно не отражается.
Календарный ход фенологических явлений у ветреницы лютичной в Подмосковье (Кожевников, 1931; Серебряков, 1947; Каляев, 1971) может быть представлен следующим образом: подснежный рост — середина февраля — середина апреля; рост надземных органов— конец апреля — конец мая; цветение — конец апреля— середина мая; созревание плодов — середина мая — начало июня; отмирание надземных органов — конец мая — середина июня; рост корневищ — май — середина июня; рост корней — июль — октябрь; формирование листовых зачатков в почках возобновления — август — сентябрь; формирование цветков — сентябрь — октябрь. По В. И. Долгошову (1947), средняя дата зацветания ветреницы лютичной в Подмосковье — 26 апреля (самое ранее зацветание 14 апреля, самое позднее — 8 мая).
По наблюдениям в лесостепи (Горышина, 1969), колебания в сроках начала и конца вегетации в годы с разными погодными условиями могут составлять около трех недель. При этом сдвиги фенодат оказываются уже, чем амплитуда колебаний погодных условий за ряд лет; следовательно, сезонный ритм носит в известной мере эндогенный характер.
Особи разных возрастных групп в популяции обнаруживают в основном сходный сезонный ритм; всходы, как и взрослые особи, имеют летний покой. Г. П. Рысина (1969) отмечает осеннее развертывание зеленого листа у проростка. В то же время весной листья проростков появляются позже, чем у взрослых растений, отмирают же они раньше (Старостенкова, 1971).
Со спецификой сезонного ритма связан ряд эколого-физиологических особенностей растения. В период надземной вегетации оно живет в условиях полной освещенности, периодически испытывает интенсивный солнечный нагрев и резкие перепады температур (ночные и утренние заморозки, разница температур между нагретым воздухом и холодной почвой днем); верхние слои почвы насыщены водой и быстро обогащаются элементами минерального питания за счет разложения подстилки (Зонн, Алешина, 1953). Таким образом, водоснабжение, почвенное и воздушное питание не лимитируют развитие растения; особенности его физиологии связаны в первую очередь с приспособлениями к температурному режиму. По данным Т. К. Горышиной, у ветреницы лютичной, как и у других ранневесенних эфемероидов, очень низок температурный порог ростовых процессов — рост может идти при любой положительной температуре. Вероятно, возможно поглощение воды даже из мерзлой почвы, хотя осмотическое давление в клетках корней невысокое. Позеленение листьев еще под снегом позволяет избежать весеннего хлороза. Цветки в закрытом состоянии без повреждения выносят даже сильные заморозки. Однако мороз около —12° в течение нескольких часов губит вегетирующее растение; в зиму с сильными морозами наблюдалась массовая гибель цветочных зачатков (Трофимов, 1949); в ходе вегетации холодостойкость растения постепенно падает.
Вегетативным органам ветреницы лютичной свойственна высокая насыщенность водой тканей и малая устойчивость к завяданию (листья могут терять не более 30—40% своего водного запаса). Транспирация очень активная, водный запас в листьях сменяется быстро (100—150% за 1 ч); интенсивность дыхания и фотосинтеза тоже высокие — приближаются к предельно возможным величинам. Ассимиляты, накопленные в течение вегетационного периода, полностью откладываются в запас, весенний рост и цветение идут за счет прошлогодних накоплений (Горышина, 1969).
Экспериментально изучена роль температурного фактора в регулировании сезонного ритма (Скрипчинский, Скрипчинский, 1961; Скрипчинский, 1969; Горышина, 1969). Летний покой у растения органический, наступает вскоре после отмирания надземных органов; он захватывает период наиболее высоких летних температур. Период действия высоких температур стимулирует начало органогенеза в почках возобновления и рост корней, рост же побегов остается заторможенным до осени — они проходят период осеннего органического покоя. Последний снимается действием пониженных осенних температур, но рост побегов уже невозможен по температурным условиям — наступает вынужденный зимний покой. К концу его осуществляется резкое снижение температурного порога ростовых процессов, становится возможным подснежный рост.
Способы размножения и распространения
Несмотря на непродолжительность цветения и сложные погодные условия, цветки вереницы лютичной опыляются насекомыми, самоопыления почти не наблюдается (Мельниченко, 1934; Иоффе. 1958). Опылители — некоторые жуки, мухи, трипсы, одиночные пчёлы — вылетают одновременно с цветением ветреницы и в дневные часы, когда открыты ее цветки, бывают достаточно активными; трипсы нередко ночуют в закрытых цветках. Плодоношение бывает ежегодным, завязывается до 70% плодиков (Мельниченко, 1934). Точных данных об эффективности семенного размножения в разных условиях нет, однако всходы обнаруживаются часто (Смирнова, 1968; Рысина, 1973). Нами в подмосковных лесах они находились ежегодно (1964—-1970 гг.) и не наблюдалось изменения численности по годам. Развиваться во взрослые особи всходы могут как внутри заросли своего вида, так и вне ее на рыхлой, покрытой лиственным опадом почве (гибнут в местах с очень мощной подстилкой или с моховым покровом).
Вегетативное размножение в популяциях ветреницы лютичной идет постоянно в связи с нарастанием и распадом корневищ (см. выше) и приводит к образованию зарослей (пятен, латок) в травяном покрове. Интенсивность разрастания и вегетативного размножения тем выше, чем больше в популяции мощных генеративных особей.
Фитоценология
По значению своему в жизни фитоценоза ветреница лютичная может быть отнесена к сезонным доминантам. В широколиственных лесах и производных от них вторичных сообществах она нередко господствует в весенней синузии травянистых растений. Роль этой синузии в накоплении фитомассы сообщества и в создании специфических почвенных условий, несмотря на кратковременность вегетации, вероятно, значительна и сопоставима с ролью видов летней вегетации (Карписонова, 1967; Горышина, 1969). Встречаясь в разных типах насаждений, ветреница лютичная чутко реагирует на состав травяного яруса. Так, выявлена ее отрицательная сопряженность с осокой волосистой (Митина, 1965) и злаками (Карписонова, 1961); хорошо развивается там, где в покрове преобладают сныть (Aegopodium podagraria L.) и зеленчук (Galeobdolon luteum Huds.).
Возрастной состав и структура популяций ветреницы лютичной в различных сообществах изучены мало. По нашим ориентировочным наблюдениям, нормальные и наиболее устойчивые ее популяции характерны для средневозрастных ненарушенных насаждений широколиственного леса с полнотой 0,6—0,7. Семенное размножение ее здесь всегда идет хорошо, вегетативное размножение обеспечивается средневозрастными генеративными особями, «отпада» мало. В спелых и перестойных насаждениях с разреженным древесным пологом и хорошо развитым подлеском возрастной спектр популяции сдвигается в сторону преобладания генеративных особей, но много всходов и ювенилов. При нарушении подлеска и травяного покрова (пастьба скота, вытаптывание) семенное возобновление подавляется, преобладают стареющие генеративные и старые вегетативные особи, много «отпада». В сильно нарушенных, разрушающихся насаждениях можно находить заросли ветреницы лютичной, состоящие только из мелких нецветущих особей и «отпада»; семенное размножение здесь отсутствует, прирост корневищ и их подвижность ничтожны [8].