

7.4.5. Мозжечок
Строение и функции головного мозга значительно сложнее спинного мозга. Рассмотрим, например, микроструктуру мозжечка.
Мозжечок выполняет важную функцию координации движений, регуляции и коррекции движений в ходе их выполнения без участия сознания. Мозжечок состоит из срединно-расположенной части (червя), обеспечивающей координацию движений туловища, и двух боковых полушарий, которые управляют координацией движений верхних и нижних конечностей (рис. 7–12). Внутри вещества червя и полушарий имеются ядра серого вещества.
Поверхность всего мозжечка покрыта корковым слоем серого вещества – корой мозжечка, которая выполняет наиболее сложную задачу – расшифровку поступающей разнородной информации о положении тела и его частей в пространстве и разработку оптимального решения, обеспечивающего равновесие тела в покое и при движении. Поверхность коры мозжечка очень велика, благодаря множеству складок (листков), имеющих одинаковую микроструктуру. Упорядоченное расположение нервных элементов в каждом листке напоминает структуру кристалла (рис. 7–13).
На рис. 7–13 представлена упрощенная схема одной складки (листка) структуры коры мозжечка. Слой грушевидных клеток Пуркинье (СП) создает регулярные дендритные ветвления в наружном молекулярном слое (МС) коры подобно четко спланированному саду. Сами клетки расположены правильными рядами как поперек, так и вдоль листка. К каждой клетке Пуркинье подходит одно лиановидное волокно (ЛВ). Каждое ЛВ дает ответвления к 10–15 клеткам Пуркинье. Лиановидное волокно обвивается вокруг дендритного дерева клетки и формирует множество возбуждающих синапсов, что обеспечивает исключительную надежность синаптической передачи.
Под слоем клеток Пуркинье находится гранулярный (зернистый) слой (ГС) с многочисленными клетками-зернами (КЗ) (более 2∙106 в 1мм3). Мшистые волокна (МВ) образуют синаптические контакты с дендритами клеток-зерен. Мшистые волокна имеют обширное ветвление (дивергенцию, от позднелат. divergentia – расхождение) связей, что обеспечивает возбуждение ими большого количества клеток в коре мозжечка.
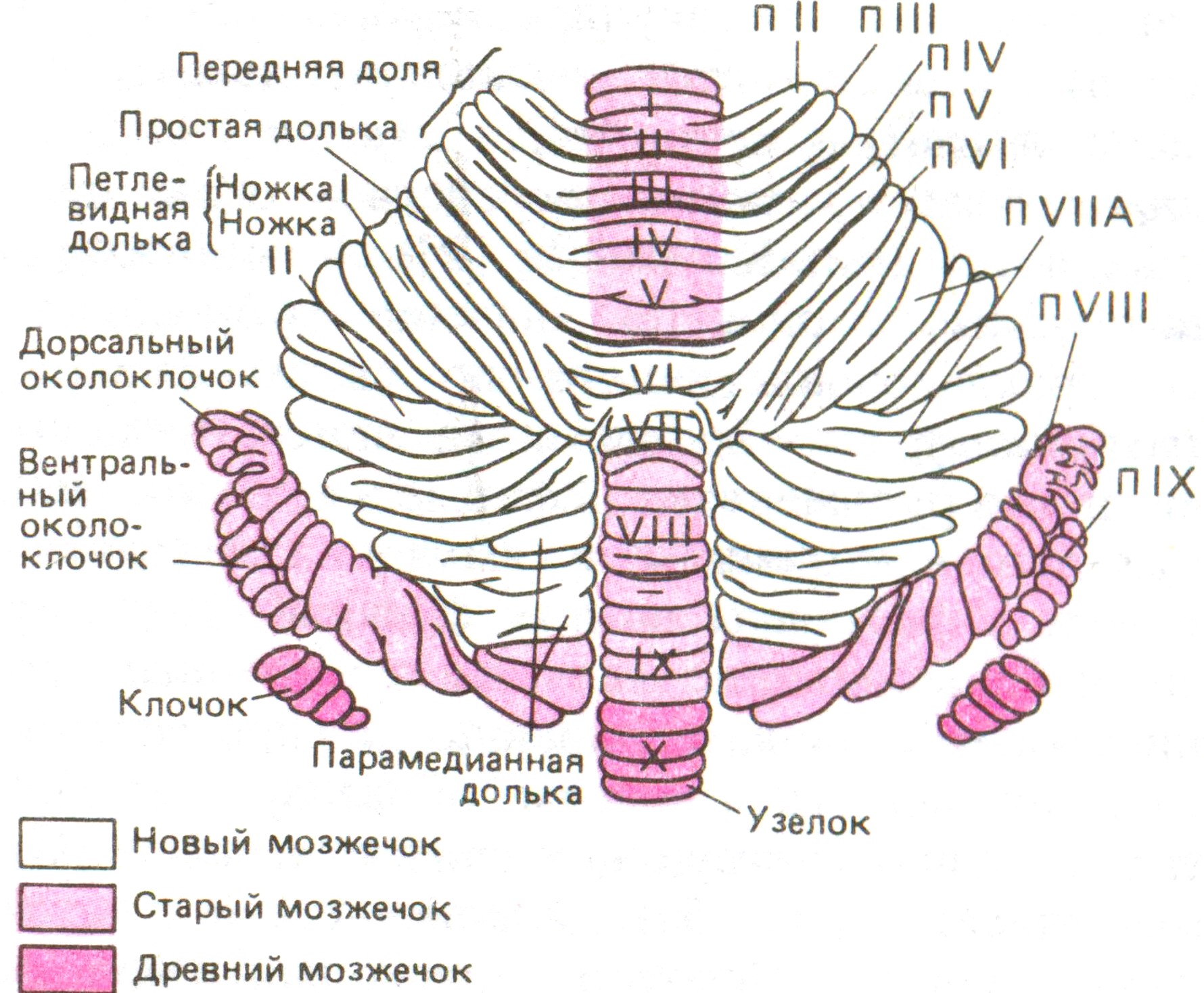
Рис. 7–12. Схема областей коры мозжечка приматов (обезьяны). Передняя доля представлена в той же плоскости, что и задняя доля. Дольки обозначены римскими цифрами (I-IX), а соответствующие части полушарий символами ПII-ПIX. У коры характерная складчатая (листовая) структура, образованная множеством листков (срез листка см. на рис. 7–13).
Аксоны клеток-зерен Т-образно разделяются на два параллельных волокна (ПВ), идущих вдоль листка в противоположных направлениях через молекулярный слой (МС) слой дендритных ветвлений. Подсчитано, что через дендритное дерево каждой клетки Пуркинье проходят под прямым углом к нему свыше 200 000 параллельных волокон. Благодаря таким волокнам на клетках коры мозжечка происходит конвергенция (от лат. converge — приближаюсь, схожусь, схождение) огромного количества мшистых волокон.
Клетки Пуркинье обеспечивают выход сигналов из коры мозжечка. Они оказывают тормозное действие на нейроны-мишени ядер мозжечка и ядра вестибулярного аппарата, обеспечивающего ориентацию головы и положения тела в пространстве. Усиление активности клеток Пуркинье относительно фонового уровня, обусловленное возбуждающими сигналами мшистых (МВ) и лиановидных (ЛВ) волокон, углубляет торможение. Под влиянием тормозных интернейронов (клеток Гольджи, корзинчатых и звездчатых клеток) происходит ослабление активности клеток Пуркинье и, как следствие, расторможение нейронов-мишеней.
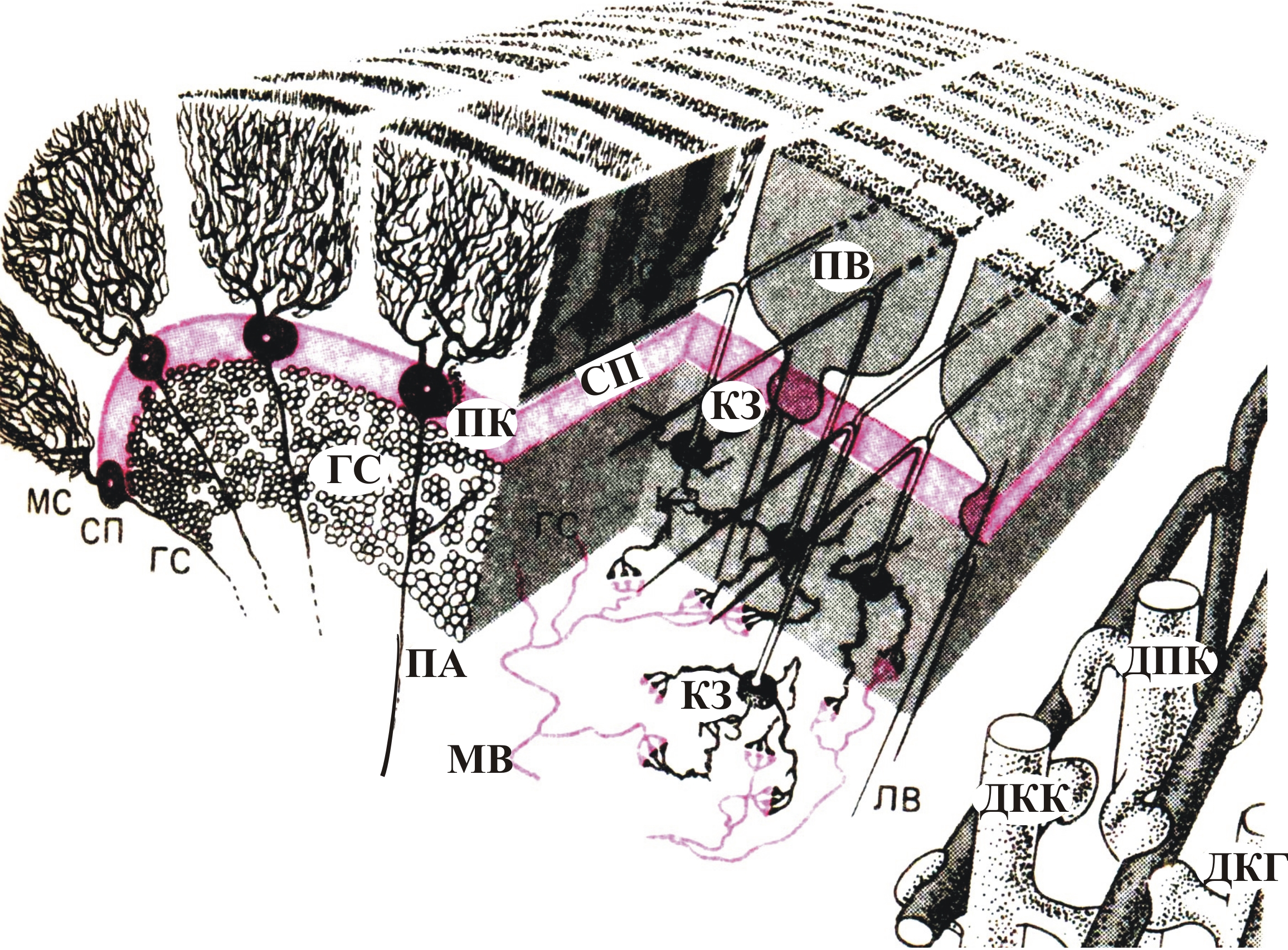
Рис. 7–13. Упрощенная схема слоистой структуры листка (поперечный срез через листок) коры мозжечка: МС – наружный молекулярный слой, СП – слой клеток Пуркинье, ГС – гранулярный слой. Клетки Пуркинье (ПК) обладают мощными дендритными ветвлениями и являются единственными выходными элементами коры мозжечка: от них в белое вещество мозжечка идет аксон (ПА). Клетки-зерна (КЗ) имеют синаптические входы от мшистых волокон (МВ) и Т-образные аксоны, которые идут вдоль листка в виде параллельных волокон (ПВ). Синаптические контакты параллельных волокон аксонов клеток-зерен с дендритами клеток Гольджи (ДКГ), дендритами корзинчатых клеток (ДКК) и дендритами клеток Пуркинье (ДКП) показаны в увеличенном виде внизу справа. Входные сигналы поступают как через мшистые волокна (МВ), так и через лиановидные волокна (ЛВ). Корзинчатые клетки, клетки Гольджи и звездчатые клетки представляют собой местные интернейроны (вставочные, промежуточные)
Существует гипотеза, согласно которой мозжечок является функциональным ответвлением главной оси «кора больших полушарий – спинной мозг». Мозжечок получает одновременно копию афферентации (информации о текущем состоянии, от лат. afferens – приносящий) и копию эфферентации (представление о требуемом конечном состоянии, от лат. efferens – выносящий) от двигательных центров (рис. 7–14). Сопоставляя первое и второе, кора мозжечка рассчитывает ошибку, о которой сообщает в двигательные центры через свои выходные ядра. Таким образом, мозжечок непрерывно корректирует и преднамеренные и автоматические движения.
Чрезвычайно важную роль в развитии мозга играют стадии его формирования в эмбриональный и постэмбриональный периоды. Поскольку нейроны не делятся, то их количество в мозге к моменту рождения определяется условиями развития мозга в эмбриональный период. Процессы,
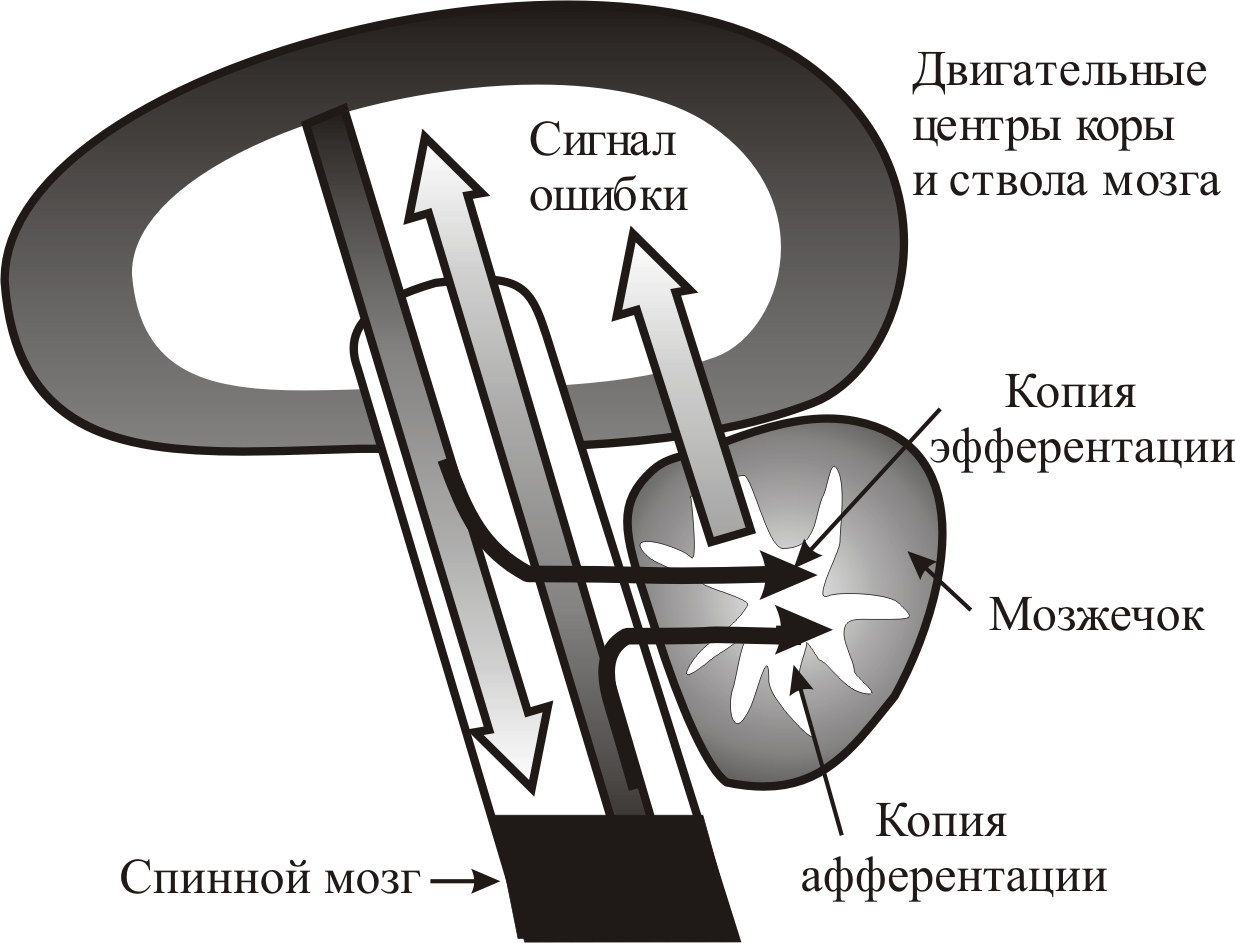
Рис. 7–14. Упрощенная схема гипотезы о сопоставительной функции мозжечка. По восходящим путям (длинная широкая стрелка вверх) от рецепторов идет афферентная информация. Двигательные центры по нисходящим в спинной мозг путям (длинная широкая стрелка вниз) посылают соответствующие эфферентные команды. Мозжечок получает копии афферентации и эфферентации (черные стрелки), сравнивая которые посылает сигнал ошибки (короткая широкая стрелка вверх) в двигательные центры, осуществляя непрерывную коррекцию двигательной программы
нарушающие нормальный синтез нейронов, делают мозг необратимо «малонейронным». При этом неизбежно нарушается нормальное формирование связей между нейронами (образование синапсов). Отрицательное действие на нейроны в эмбриональный период развития оказывают алкоголь, никотин, наркотики. В результате их действия возникают отклонения от нормы и у вспомогательных клеток – глиоцитов, и у кровеносных сосудов.
Обратим внимание на то, что масса мозга в эмбриональном периоде у (15-26) недельных эмбрионов составляет приблизительно 16% от массы тела, при рождении ~10%, а у взрослого человека – менее 2%. Уже в эмбриональном периоде головной мозг участвует в регуляции развития эндокринных желез, реагирует на тактильные (от лат. tactilis – осязаемый), звуковые, вкусовые раздражители, формирует рефлекторные реакции.
Масса мозга быстро увеличивается во времени. У новорожденных она составляет 350-380 г, у годовалого ребенка ~1 000 г, к 5–6 годам масса мозга у ребенка приближается к массе мозга взрослого человека: в среднем 1375 г у мужчин и 1275 г у женщин.
Таким образом, генетическая программа, определяющая свойства головного мозга, может реализоваться в полном объеме только при создании оптимальных условий для развития эмбриона.
Одной из специфических особенностей мозга человека, в отличие от мозга животных, является то, что уже у новорожденных функционирует механизм восприятия различных признаков речевых сигналов, то есть человеческий мозг обладает врожденной способностью освоения языка. Именно на основе врожденных механизмов под влиянием внешней среды у ребенка формируется восприятие речи.
Сходство физических характеристик акустических сигналов у новорожденных детей разных национальностей указывает на врожденный характер способности генерировать речевые звуки. Только с шести месяцев у младенцев начинают проявляться особенности генерируемых звуков, характерные для языкового окружения, исчезают звуки нехарактерные для родного языка.