

 л.
VI. Физико-химические
процессы в биологических мембранах
л.
VI. Физико-химические
процессы в биологических мембранах
VI. Физико-химические процессы в биологических мембранах §6.1. Структура мембран
Биологические мембраны – сложные высокоорганизованные надмолекулярные структуры, ограничивающие клетки (клеточные или плазматические мембраны) и внутриклеточные органоиды – митохондрии, лизосомы и т.п. Мембраны представляют собой пленки толщиной (5 10) нм, состоящие, главным образом, из белков и липидов (рис. 6–1). Общая площадь мембран в организме человека составляет десятки тысяч квадратных метров.
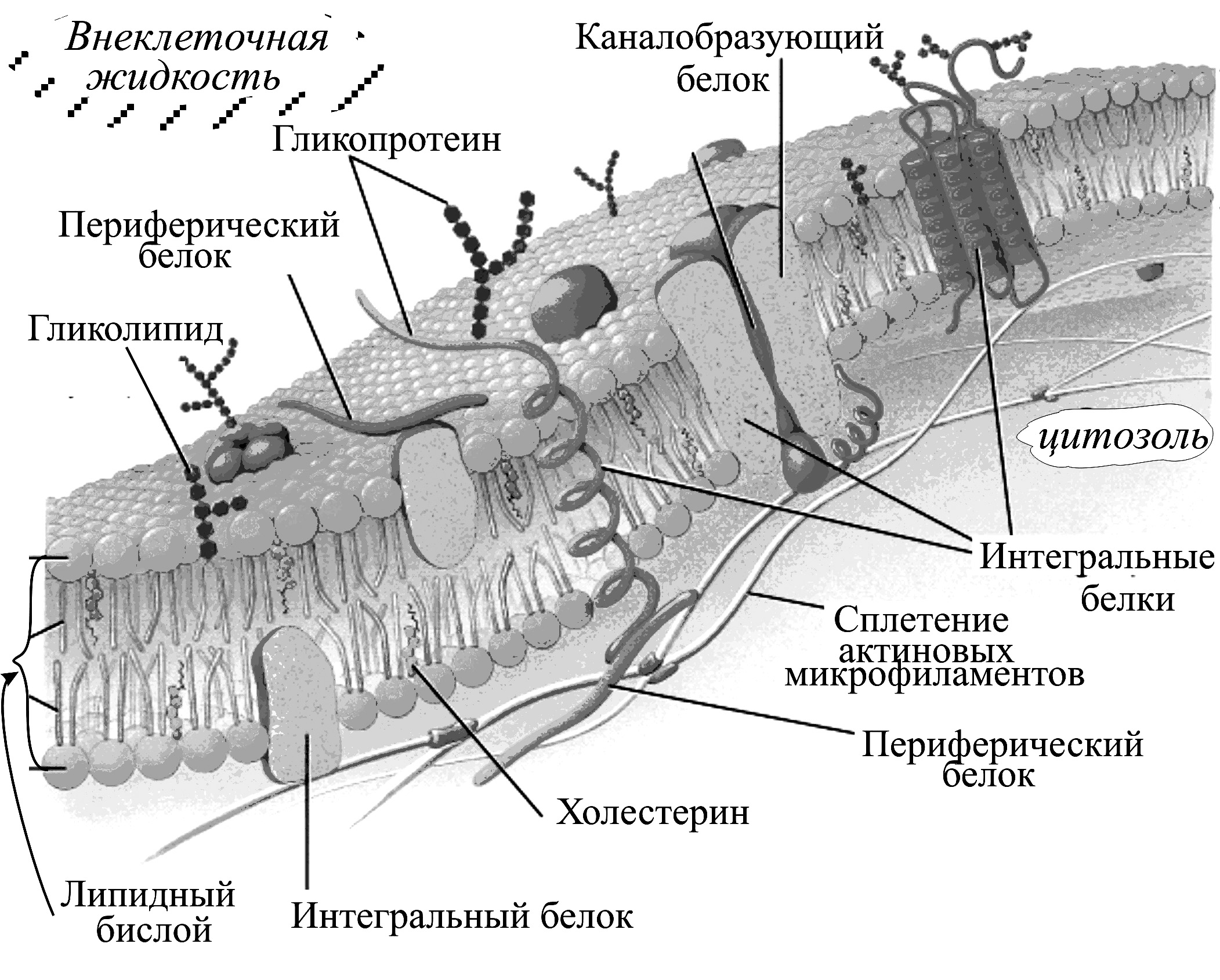
Рис. 6–1. Структура плазматической мембраны, состоящей из двух слоев липидных молекул (липидный бислой), в которые встроены белки. Углеводы связаны с белками (гликопротеины) и липидами (гликолипиды). Кроме фосфолипидов (главной составляющей липидного бислоя) и гликолипидов важную группу составляет третий класс липидов – холестерин, находящийся внутри мембраны, который способен легко переходить с одной стороны мембраны на другую. Мембранные белки разделяются на интегральные – трансмембранные, спирализованные участки которых одно- или многократно пересекают липидный бислой, и периферические белки, удерживаемые на мембране с помощью липидного «якоря». Расположенные под плазматической мембраной сплетенные актиновые нити микрофиламентов участвуют в изменении формы клетки
Основные липидные компоненты мембран – фосфолипиды, гликолипиды и стерины (см. с. 385–392), плоские кольца которых ограничивают подвижность мембран и придают мембранам жесткость. Важным компонентом животных мембран является холестерин.
6.1.1. Жидкокристаллическая модель мембран
В 1972 г. была предложена жидкокристаллическая (ч.III, с. 155–169) модель строения мембран. По этой модели, структурным остовом мембраны является фосфолипидный бислой (рис. 6–1).
Молекула фосфолипида (см. рис. 3–19, 3–21, с. 389-390) имеет длинные гидрофобные фрагменты углеводородной цепи в виде незаряженных "хвостов" и небольшой полярный гидрофильный фрагмент ("головку").
В клетке по обе стороны клеточной мембраны находятся водные фазы электролита – наружный раствор и клеточное содержимое. Наиболее энергетически выгодным для фосфолипидов является образование плоского бимолекулярного слоя (кластера) толщиной порядка (4 5) нм. В нем гидрофильные части фосфолипидных молекул направлены к водным фазам, а их гидрофобные части – внутрь мембраны. Распределение различных липидов во внутреннем и наружном слоях мембраны неодинаково. Более того, даже в пределах одного слоя имеются участки, в которых концентрируются отдельные виды липидов. Функциональное значение такого неравномерного распределения различных сортов липидов остается пока неясным.
Липидная мембрана имеет на обеих поверхностях заряд, который создается полярными головками фосфолипидных молекул. Этот заряд экранируется противоположно заряженными ионами окружающего мембрану электролита. Заряды формируют диффузные обкладки двойного электрического слоя. В результате образуется поверхностный скачок потенциала, величина которого зависит от плотности поверхностного заряда на мембране и концентрации электролита.
Мембрана является
очень хорошим изолятором, обладая
высоким электрическим сопротивлением
![]() .
Концентрации ионов в водной и липидной
фазах отличаются в ~1012 раз.
Диэлектрическая проницаемость гидрофобной
сердцевины мембран равна ~2, то есть в
~40 раз меньше, чем у воды. Биологическую
мембрану можно рассматривать как
конденсатор с удельной емкостью на
единицу площади
.
Концентрации ионов в водной и липидной
фазах отличаются в ~1012 раз.
Диэлектрическая проницаемость гидрофобной
сердцевины мембран равна ~2, то есть в
~40 раз меньше, чем у воды. Биологическую
мембрану можно рассматривать как
конденсатор с удельной емкостью на
единицу площади
![]() .
Роль пластин конденсатора выполняют
электролиты цитоплазмы и внеклеточного
раствора. Используя соотношение для
емкости плоского конденсатора
.
Роль пластин конденсатора выполняют
электролиты цитоплазмы и внеклеточного
раствора. Используя соотношение для
емкости плоского конденсатора
![]() ,
можно оценить толщину мембраны
,
можно оценить толщину мембраны
![]() (при площади пластин
(при площади пластин
![]() ).
).
При физиологических температурах бислой находится в жидкокристаллическом состоянии (см. с. 155-169): в перпендикулярном направлении он ведет себя как твердое тело, а в боковом (латеральном, от лат. lateralis – боковой), вдоль плоскости – как липидная жидкость, инкрустированная белками. Такое анизотропное строение мембраны, с одной стороны, позволяет защищать ее внутреннюю часть от нежелательных внешних воздействий, а с другой стороны, ее жидкостный характер обеспечивает высокие транспортные свойства (проницаемость, перенос ионов и др.).
При понижении температуры фосфолипидные мембраны испытывают фазовый переход первого рода из жидкокристаллического состояния в гель–состояние (рис. 6–2).

Рис. 6–2. Изменение структуры мембраны из лецитина при переходе из жидкокристаллического в гель (твердокристаллическое) состояние и обратно
Гель–состояние (т.1, с.175) является более упорядоченным, чем жидкокристаллическое. Все гидрофобные углеводородные «хвосты» фосфолипидных молекул в гель–фазе полностью вытянуты. Толщина мембраны в гель–фазе больше, чем в жидкокристаллическом состоянии. Однако объем при переходе в гель–состояние уменьшается в результате преобладающего уменьшения площади мембраны, приходящейся на одну молекулу (от (7,6Å)2=0,58 нм до (6,9Å)2=0,48 нм).
В зависимости от химического состава липидных мембран температура фазового перехода варьируется от –20С (для мембран ненасыщенных липидов) до +60С (в случае насыщенных липидов).
Для нормального функционирования мембрана должна быть в жидкокристаллическом состоянии. Поэтому у организмов, существующих в низкотемпературной среде, например, в ногах у северного оленя, клеточные мембраны содержат больше ненасыщенных липидов, чем в остальных клетках тела. В связи с этим можно предположить, что первичный механизм криоповреждений (при охлаждении) связан с переходом мембран в гель–состояние.
Наличие большого количества холестерина в мембранах можно объяснить тем, что он смягчает (уменьшает) изменения в мембране при фазовом переходе. Молекулы холестерина, располагаясь между фосфолипидными молекулами, упорядочивают бислой в жидкокристаллическом состоянии и разупорядочивают – в гель-состоянии.
Обычно мембраны содержат много различных липидов, отличающихся температурами фазовых переходов, в том числе липидов, температура фазового перехода которых близка к физиологической.