

5.10.4. Модели индуктивно-резонансной миграции энергии
Как уже говорилось, индуктивно-резонансный перенос энергии может происходить как между молекулами, так и внутри одной молекулы. Если макромолекула имеет несколько степеней свободы, между которыми имеется связь, то возбуждение одной из степеней свободы приводит, благодаря наличию взаимодействия, к возбуждению других степеней свободы макромолекулы. Таким образом, связанные друг с другом степени свободы могут обмениваться энергией.
Механическая модель
Механическим аналогом, поясняющим
механизм индуктивно – резонансного
переноса энергии могут служить два
одинаковых маятника, связанных друг с
другом пружинкой (рис. 5–21):
![]() – собственная частота колебаний
маятников в отсутствие связи, b0
– коэффициент жесткости пружин маятников,
m – масса маятников,
b – коэффициент
жесткости пружины связи (коэффициент
связи). Условие слабой связи записывается
в виде
– собственная частота колебаний
маятников в отсутствие связи, b0
– коэффициент жесткости пружин маятников,
m – масса маятников,
b – коэффициент
жесткости пружины связи (коэффициент
связи). Условие слабой связи записывается
в виде
![]() . (5.27)
. (5.27)
Уравнения движения маятников имеют вид
![]() , (5.28)
, (5.28)
![]() , (5.29)
, (5.29)
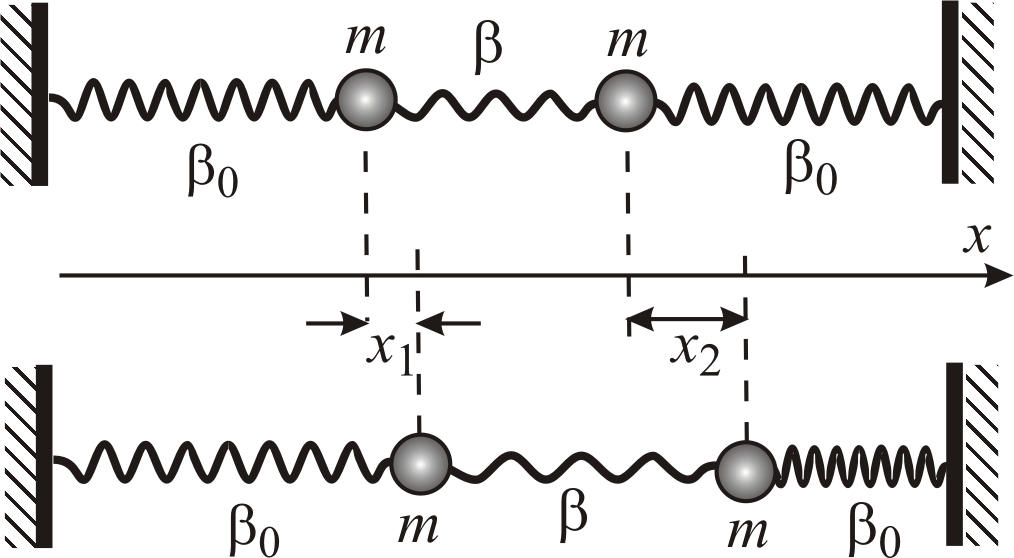
где х1 и х2 – смещения из положений равновесия, соответственно, первого и второго маятников (рис. 5–21).
Деля на массу и складывая уравнения (5.28) и (5.29), получаем
![]() , (5.30)
, (5.30)
а вычитая –
![]() . (5.31)
. (5.31)
|
Рис. 5–21. Система
двух связанных маятников с сосредоточенными
параметрами (шарики с массой m
не обладают упругостью, пружины с
коэффициентами жесткости
и
|
Уравнения (5.30) и (5.31) представляют собой уравнения гармонических колебаний (нормальных колебаний системы связанных маятников). Общее решение уравнения (5.30) записывается в виде
![]() (5.32)
(5.32)
где частота
![]() .
(5.33)
.
(5.33)
Решение уравненияе (5.31) имеет вид
![]() ,
(5.34)
,
(5.34)
где частота w2 с учетом (5.27) приближенно равна
![]() (5.35)
(5.35)
![]() (5.36)
(5.36)
Амплитуды С1, С2 и начальные фазы φ1, φ2 определяются начальными условиями (смещениями из положений равновесия и скоростями при t = 0). Зададим начальные условия в момент времени t = 0:
![]() ,
,
![]() ,
,
![]() , (5.37)
, (5.37)
т. е. первому маятнику сообщена кинетическая
энергия
![]() .
.
Начальные условия (5.37) определяют фазы
и амплитуды колебаний:
![]() и
и
![]() ,
,
![]() .
Уравнения (5.32) и (5.33) принимают вид:
.
Уравнения (5.32) и (5.33) принимают вид:
![]() ,
(5.38)
,
(5.38)
![]() ,
(5.39)
,
(5.39)
Из (5.38) и (5.39) получаем законы движения маятников
![]() , (5.40)
, (5.40)
![]() . (5.41)
. (5.41)
и, используя, выражения для их Кинетическая энергия движения маятников, с учетом (5.35), описывается выражениями (рис. 5–22):
![]()
![]() ,
(5.42)
,
(5.42)
![]() .
(5.43)
.
(5.43)
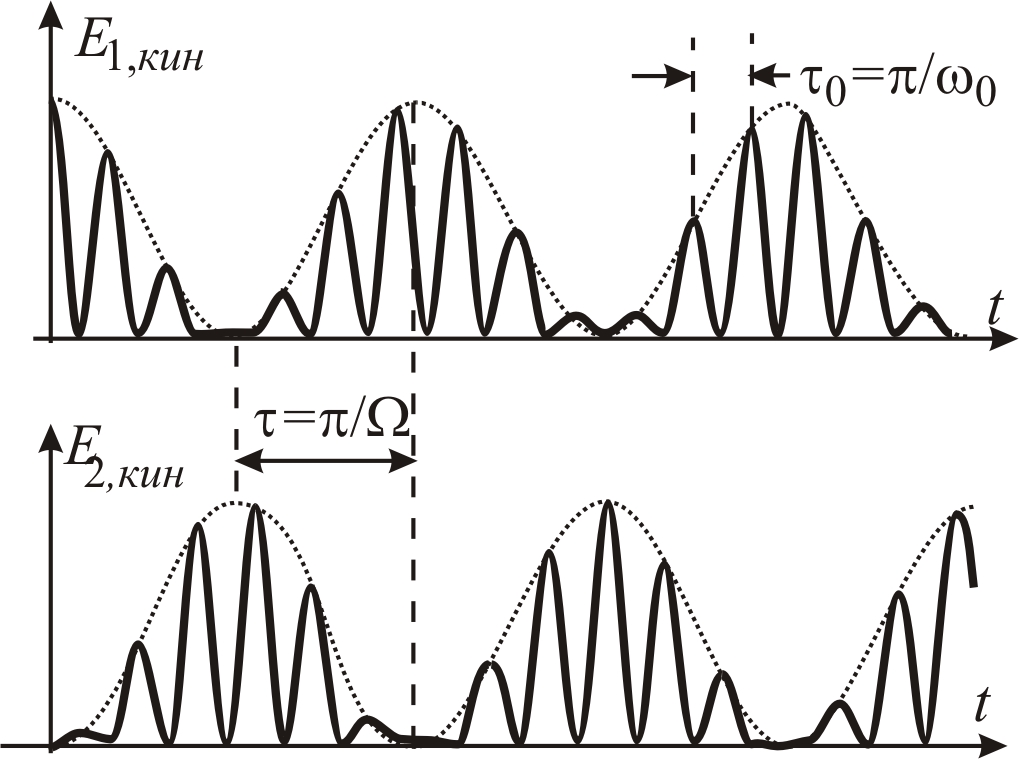
Рис. 5–22.
Зависимости от времени кинетических
энергий
![]() и
и
![]() первого и второго маятников;
первого и второго маятников;
![]() — период изменения полной энергии
каждого маятника (время перехода энергии
из кинетической в потенциальную и
обратно);
— период изменения полной энергии
каждого маятника (время перехода энергии
из кинетической в потенциальную и
обратно);
![]() – время перекачки энергии от одного
маятника к другому
– время перекачки энергии от одного
маятника к другому
Из рис. 5–22 видно, что, во-первых,
кинетическая энергия каждого маятника
периодически с частотой 2w0
(периодом
![]() )
переходит из кинетической в потенциальную
и обратно и, во-вторых, – через время
)
переходит из кинетической в потенциальную
и обратно и, во-вторых, – через время
![]() происходит перекачка энергии от одного
маятника к другому. Процесс обмена
энергиями между маятниками полностью
обратим. Если в рассматриваемой системе
учесть возможную диссипацию энергии,
то обмен энергиями будет происходить
с частотой до тех пор, пока колебания
не затухнут.
происходит перекачка энергии от одного
маятника к другому. Процесс обмена
энергиями между маятниками полностью
обратим. Если в рассматриваемой системе
учесть возможную диссипацию энергии,
то обмен энергиями будет происходить
с частотой до тех пор, пока колебания
не затухнут.
Аналогичный результат можно получить для двух связанных молекулярных осцилляторов. Поскольку процесс переноса энергии между двумя одинаковыми молекулами в стационарных условиях является обратимым, то он не приводит к направленному переносу энергии, а может приводить лишь к ее миграции от одной молекулы к другой.